 Ulaz
UlazTubularna reapsorpcija i transport esencijalnih minerala. Mehanizmi reapsorpcije u tubulima
tekstualna_polja
tekstualna_polja
arrow_upward
Poređenje sastava i količine primarnog i konačnog urina pokazuje da u tubulima nefrona dolazi do procesa reapsorpcije vode i supstanci filtriranih u glomerulima. Ovaj proces se zove tubularna reapsorpcija
U zavisnosti od odjeljenja tubula gdje se javlja, postoje reapsorpcija proksimalni idistalno.
Reapsorpcija je transport tvari iz urina u limfu i krv a zavisno od mehanizma transporta izoluju se pasivna, primarna i sekundarna aktivna reapsorpcija.
proksimalna reapsorpcija
tekstualna_polja
tekstualna_polja
arrow_upward
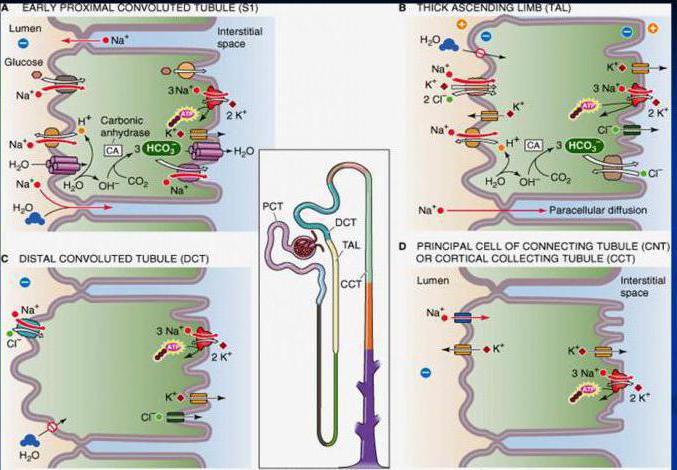
Proksimalna reapsorpcija osigurava potpunu apsorpciju niza primarnih supstanci urina - glukoze, proteina, aminokiselina i vitamina. U proksimalnim dijelovima se apsorbira 2/3 filtrirane vode i natrijuma, velike količine kalija, dvovalentnih katjona, hlora, bikarbonata, fosfata, kao i mokraćne kiseline i uree. Do kraja proksimalnog presjeka u njegovom lumenu ostaje samo 1/3 volumena ultrafiltrata, a iako se njegov sastav već značajno razlikuje od krvne plazme, osmotski tlak primarnog urina ostaje isti kao u plazmi.
Usisavanje vode javlja se pasivno, duž gradijenta osmotskog pritiska i zavisi od reapsorpcije natrijuma i hlorida. Reapsorpcija natrijum u proksimalnom dijelu vrši se i aktivnim i pasivnim transportom. U početnom dijelu tubula, ovo je aktivan proces. Iako natrijum ulazi u epitelne ćelije kroz apikalnu membranu pasivno kroz natrijeve kanale duž koncentracijskog i elektrohemijskog gradijenta, njegovo izlučivanje kroz bazolateralne membrane epitelnih ćelija odvija se aktivno uz pomoć natrijum-kalijum pumpi koristeći ATP energiju. Popratni apsorbovani natrijum anjon je ovde bikarbonat, a hloridi se slabo apsorbuju. Volumen urina u tubulu se smanjuje zbog pasivne reapsorpcije vode, a koncentracija klorida u njegovom sadržaju se povećava. U terminalnim dijelovima proksimalnih tubula, međućelijski kontakti su visoko propusni za kloride (čija je koncentracija povećana) i pasivno se apsorbiraju iz urina duž gradijenta. Zajedno s njima, natrijum i voda se pasivno reapsorbuju. Takav pasivni transport jednog jona (natrijuma) zajedno sa pasivnim transportom drugog (hlorida) naziva se cotransport.
Dakle, u proksimalnom nefronu postoje dva mehanizma za apsorpciju vode i jona:
1) aktivni transport natrijuma uz pasivnu reapsorpciju bikarbonata i vode,
2) pasivni transport hlorida uz pasivnu reapsorpciju natrijuma i vode.
Pošto se natrijum i drugi elektroliti uvek apsorbuju u proksimalne tubule sa osmotski ekvivalentnom količinom vode, urin u proksimalnom nefronu ostaje izosmotičan krvnoj plazmi.
proksimalna reapsorpcija glukoze i amino kiseline provodi se uz pomoć posebnih nosača četkice apikalne membrane epitelnih stanica. Ovi transporteri prenose glukozu ili aminokiseline samo ako se vežu i transportuju natrijum. Pasivno kretanje natrijuma duž gradijenta u ćelije dovodi do prolaska kroz membranu i nosač sa glukozom ili aminokiselinom. Za implementaciju ovog procesa potrebna je niska koncentracija natrijuma u ćeliji, što stvara koncentracijski gradijent između vanjskog i intracelularnog okruženja, što se osigurava energetski ovisnim radom natrijum-kalijum pumpe bazalne membrane. Budući da je prijenos glukoze ili aminokiseline povezan s natrijumom, a njegov transport je određen aktivnim uklanjanjem natrijuma iz ćelije, ova vrsta transporta se naziva sekundarno aktivan ili simport, one. zajednički pasivni transport jedne supstance (glukoze) zbog aktivnog transporta druge (natrijuma) pomoću jednog nosača.
Budući da je za reapsorpciju glukoze potrebno svaki njen molekul vezati za molekulu nosača, očito je da se uz višak glukoze svi molekuli nosači mogu potpuno opteretiti i glukoza se više ne može apsorbirati u krv. Ovu situaciju karakteriše "maksimalni tubularni transluka materije",što odražava maksimalno opterećenje tubularnih nosača pri određenoj koncentraciji tvari u primarnom urinu i, prema tome, u krvi. Postepeno povećavajući sadržaj glukoze u krvi, a time i u primarnom urinu, lako se može pronaći vrijednost njene koncentracije pri kojoj se glukoza pojavljuje u konačnom urinu i kada njeno izlučivanje počinje linearno ovisiti o porastu razine u krvi. Ova koncentracija glukoze u krvi i, shodno tome, u ultrafiltratu ukazuje da su svi tubularni transporteri dostigli granicu funkcionalnosti i da su potpuno opterećeni. U ovom trenutku, reapsorpcija glukoze je maksimalna i kreće se od 303 mg/min kod žena do 375 mg/min kod muškaraca. Vrijednost maksimalnog cjevastog transporta odgovara starijem konceptu "bubrežniprag povlačenja.
Prag bubrežne eliminacije naziva se koncentracija tvari u krvi i primarnom urinu pri kojoj se više ne može potpuno reapsorbirati u tubulima i pojavljuje se u konačnom urinu.
Takve supstance za koje se može pronaći prag eliminacije, tj. nazivaju se potpuno reapsorbirani pri niskim koncentracijama u krvi, a ne u potpunosti u povišenim koncentracijama prag. Tipičan primjer je glukoza, koja se potpuno apsorbira iz primarnog urina pri koncentracijama u plazmi ispod 10 mol/l, ali se pojavljuje u konačnom urinu, tj. ne resorbuje se u potpunosti, kada je njegov sadržaj u krvnoj plazmi iznad 10 mol/l. Stoga je za glukozu prag eliminacije 10 mol/l.
Tvari koje se uopće ne reapsorbiraju u tubulima (inulin, manitol) ili se malo reapsorbiraju i izlučuju srazmjerno nakupljanju u krvi (urea, sulfati itd.) nazivaju se bez praga, jer Za njih ne postoji prag za povlačenje.
Male količine filtriranog vjeverica skoro potpuno reapsorbuje u proksimalnim tubulima pinocitozom. Mali proteinski molekuli apsorbiraju se na površini apikalne membrane epitelnih stanica i apsorbiraju ih uz stvaranje vakuola, koje se pri kretanju spajaju s lizosomima. Proteolitički enzimi lizosoma razgrađuju apsorbirani protein, nakon čega se fragmenti male molekularne težine i aminokiseline prenose u krv kroz bazolateralnu membranu stanica.
Distalna reapsorpcija
tekstualna_polja
tekstualna_polja
arrow_upward
Distalna reapsorpcija jona i vode je po zapremini mnogo manja od proksimalne. Međutim, značajno mijenjajući se pod utjecajem regulatornih utjecaja, on određuje sastav konačnog urina i sposobnost bubrega da izlučuje koncentrirani ili razrijeđeni urin (u zavisnosti od ravnoteže vode u tijelu). Aktivna reapsorpcija se javlja u distalnom nefronu ontria. Iako se ovdje apsorbira samo 10% filtrirane količine kationa, ovaj proces osigurava izraženo smanjenje njegove koncentracije u urinu i, obrnuto, povećanje koncentracije u intersticijskoj tekućini, što stvara značajan gradijent osmotskog tlaka između urina. i intersticijum. Hlor apsorbuje se uglavnom pasivno nakon natrijuma. Sposobnost epitela distalnih tubula da luči H-jone u mokraću povezana je sa reapsorpcijom natrijumovih jona, ova vrsta transporta u vidu razmene natrijuma za proton se naziva "antiport". Aktivno se apsorbira u distalnim tubulima kalijum, kalcijum i phosvelovi. U sabirnim kanalima, uglavnom jukstamedularnim nefronima, pod uticajem vazopresina, permeabilnost zida na urea a zbog visoke koncentracije u lumenu tubula pasivno difundira u okolni intersticijski prostor, povećavajući njegovu osmolarnost. Pod uticajem vazopresina, zid distalnih uvijenih tubula i sabirnih kanala postaje propustljiv za voda, kao rezultat, reapsorbuje se duž osmotskog gradijenta u hiperosmolarni intersticij medule i dalje u krv.
Sposobnost bubrega da formira koncentrirani ili razrijeđeni urin osigurava aktivnost protivstrujno umnožavatitelesni cevasti sistem bubreg, koji je predstavljen paralelnim kolenima Henleove petlje i sabirnim kanalićima (slika 12.2).
Brojevi označavaju vrijednosti osmotskog tlaka intersticijske tekućine i urina. U sabirnom kanalu brojevi u zagradi označavaju osmotski pritisak urina u odsustvu vazopresina (razblaživanje urina), brojevi bez zagrada označavaju osmotski pritisak urina pod dejstvom vazopresina (koncentracija urina).
Urin se u ovim tubulima kreće u suprotnim smjerovima (zbog čega je sistem nazvan protustrujnim), a procesi transporta supstanci u jednom koljenu sistema se pojačavaju („umnožavaju“) zbog aktivnosti drugog koljena. Uzlazno koljeno Henleove petlje igra odlučujuću ulogu u radu protustrujnog mehanizma, čiji je zid nepropustan za vodu, ali aktivno reapsorbira ione natrija u okolni intersticijski prostor. Kao rezultat, intersticijska tekućina postaje hiperosmotska u odnosu na sadržaj silaznog kraka petlje, a prema vrhu petlje raste osmotski tlak u okolnom tkivu. Zid descendentnog koljena je propustljiv za vodu, koja se pasivno kreće iz lumena u hiperosmotski intersticij. Tako u silaznom koljenu urin postaje sve više hiperosmotski zbog apsorpcije vode, tj. uspostavlja se osmotska ravnoteža sa intersticijskom tečnošću. U uzlaznom koljenu, zbog apsorpcije natrijuma, urin postaje sve manje osmotski i već hipotonični urin se penje u kortikalni dio distalnog tubula. Međutim, njegova količina, zbog apsorpcije vode i soli u Henleovoj petlji, značajno se smanjila.
Sabirni kanal, u koji zatim ulazi urin, takođe formira protivstrujni sistem sa uzlaznim ekstremitetom Henleove petlje. Zid sabirnog kanala postaje propustljiv za vodu samo u prisustvu vazopresin. U ovom slučaju, kako se urin kreće duž sabirnih kanala duboko u medulu, u kojoj se osmotski tlak povećava zbog apsorpcije natrijuma u uzlaznom dijelu Henleove petlje, sve više i više vode prolazi pasivno u hiperosmotski intersticij i urina postaje sve više. i koncentrisanije.
Pod uticajem vazopresina ostvaruje se još jedan važan mehanizam za koncentraciju urina – pasivni izlazak ureje iz sabirnih kanala u okolni intersticij. Apsorpcija vode u gornjim dijelovima sabirnih kanala dovodi do povećanja koncentracije uree u mokraći, a u njihovim najnižim dijelovima, koji se nalaze duboko u meduli, vazopresin povećava propusnost uree i ona pasivno difunduje u intersticij. , naglo povećavajući njegov osmotski pritisak. Tako intersticij medule postaje najjače osmotski u području vrha bubrežnih piramida, gdje dolazi do povećanja apsorpcije vode iz lumena tubula u intersticij i koncentracije urina.
Urea intersticijske tekućine difundira duž gradijenta koncentracije u lumen tankog uzlaznog dijela Henleove petlje i ponovo ulazi u distalne tubule i sabirne kanale s mlazom urina. Tako se odvija cirkulacija uree u tubulima, održavajući visok nivo njene koncentracije u meduli. Opisani procesi se odvijaju uglavnom u jukstamedularnim nefronima, koji imaju najduže Henleove petlje, koje se spuštaju duboko u medulu bubrega.
U meduli bubrega nalazi se još jedan - vascular proprotivstrujni sistem, formirane od krvnih kapilara. Budući da cirkulacijska mreža jukstamedularnih nefrona formira dugačke paralelne ravne silazne i uzlazne kapilarne žile (slika 12.1), spuštajući se duboko u medulu, krv koja se kreće duž silazne direktne kapilarne žile postepeno oslobađa vodu u okolni intersticijski prostor zbog povećanja osmotski pritisak u tkivu i, obrnuto, obogaćen natrijumom i ureom, zgušnjava i usporava njegovo kretanje. U uzlaznoj kapilarnoj žili, kako se krv kreće u tkiva uz postupno opadajući osmotski tlak, dolazi do obrnutih procesa – natrijum i urea difundiraju natrag u tkivo duž gradijenta koncentracije, a voda se apsorbira u krv. Dakle, ovaj protivstrujni sistem takođe doprinosi održavanju visokog osmotskog pritiska u dubokim slojevima tkiva medule, obezbeđujući uklanjanje vode i zadržavanje natrijuma i uree u intersticijumu.
Aktivnost opisanih protustrujnih sistema u velikoj mjeri ovisi o brzini kretanja tekućina (urina ili krvi) u njima. Što prije urin krene kroz cijevi protustrujnog tubularnog sistema, to će manje natrijuma, uree i vode imati vremena da se reapsorbiraju u intersticij, a više manje koncentriranog urina će se izlučiti putem bubrega. Što je veća brzina protoka krvi kroz direktne kapilarne sudove medule bubrega, to će više natrijuma i ureje iznijeti krv iz bubrežnog intersticija, jer. neće imati vremena da difundiraju iz krvi nazad u tkivo. Ovaj efekat se zove "ispiranje" osmotski aktivne supstance iz intersticija, usled čega se smanjuje njegov osmolarnost, smanjuje se koncentracija urina i više mokraće se izlučuje preko bubrega niska specifična težina(razblaživanje urina). Što je sporije kretanje urina ili krvi u meduli bubrega, to se više osmotski aktivnih supstanci akumulira u intersticiju i veća je sposobnost bubrega koncentrat urin.
Regulacija tubularne reapsorpcije
tekstualna_polja
tekstualna_polja
arrow_upward
Regulacija tubularne reapsorpcije izvršeno kao nervozan i, u većoj mjeri, humoralni način.
Nervni uticaji se pretežno ostvaruju putem simpatičkih provodnika i medijatora preko beta-adrenergičkih receptora ćelijskih membrana proksimalnih i distalnih tubula. Simpatički efekti se manifestuju u vidu aktivacije procesa reapsorpcije glukoze, natrijuma, vode i fosfata i ostvaruju se kroz sistem sekundarnih glasnika (adenilat ciklaza - cAMP). Trofički uticaji simpatičkog nervnog sistema igraju važnu ulogu u regulaciji metaboličkih procesa u bubrežnom tkivu. Nervna regulacija cirkulacije krvi u bubrežnoj meduli povećava ili smanjuje efikasnost vaskularnog protustrujnog sistema i koncentraciju urina.
Vaskularni efekti nervne regulacije mogu biti posredovani kroz intrarenalne sisteme humoralnih regulatora - renin-angiotenzin, kinin, prostaglandin, itd. Glavni faktor u regulaciji reapsorpcije vode u distalnom nefronu je hormon vazopresin, ranije zvao antidiuretički hormon. Ovaj hormon se proizvodi u supraoptičkim i paraventrikularnim jezgrama hipotalamusa i ulazi u krvotok iz neurohipofize. Efekat vazopresina na permeabilnost tubularnog epitela je zbog prisustva hormonskih receptora tipa V-2 na površini bazolateralne membrane epitelnih ćelija. Formiranje hormonsko-receptorskog kompleksa (Poglavlje 3) podrazumijeva, preko GS-proteina i gvanil nukleotida, aktivaciju adenilat ciklaze i formiranje cAMP na bazolateralnoj membrani (slika 12.3).
Rice. 12.3. Mehanizam djelovanja vazopresina na permeabilnost sabirnih kanala za vodu.Rice. 12.3. Mehanizam djelovanja vazopresina na permeabilnost sabirnih kanala za vodu.
B-l membrana - bazolateralna membrana ćelija,
A membrana je apikalna membrana,
GN - gvanidin nukleotid, AC - adenilat ciklaza.
Nakon toga, cAMP prolazi kroz epitelnu ćeliju i, došavši do apikalne membrane, aktivira cAMP zavisne protein kinaze. Pod uticajem ovih enzima dolazi do fosforilacije membranskih proteina, što dovodi do povećanja propusnosti vode i povećanja površine membrane. Preuređenje ultrastrukture ćelije dovodi do formiranja specijalizovanih vakuola koje nose velike tokove vode duž osmotskog gradijenta od apikalne do bazolateralne membrane, sprečavajući samu ćeliju da nabubri. Takav transport vode kroz epitelne ćelije ostvaruje se vazopresinom u sabirnim kanalićima. Osim toga, u distalnim tubulima vazopresin uzrokuje aktivaciju i oslobađanje hijaluronidaza iz stanica, uzrokujući razgradnju glikozaminoglikana glavne međućelijske tvari i međućelijski pasivni transport vode duž osmotskog gradijenta.
tubularna reapsorpcija vode
tekstualna_polja
tekstualna_polja
arrow_upward
Tubularna reapsorpcija vode je također regulirana drugim hormonima.
Uzimajući u obzir mehanizme djelovanja, svi hormoni koji reguliraju reapsorpciju vode mogu se predstaviti u šest grupa:
1) povećanje permeabilnosti membrana distalnog nefrona za vodu (vazopresin, prolaktin, korionski gonadotropin);
2) promena osetljivosti ćelijskih receptora na vazopresin (paratirin, kalcitonin, kalcitriol, prostaglandini, aldosteron);
3) promena osmotskog gradijenta intersticijuma medule bubrega i, shodno tome, pasivnog osmotskog transporta vode (paratirin, kalcitriol, tiroidni hormoni, insulin, vazopresin);
4) promena aktivnog transporta natrijuma i hlorida, a zbog toga i pasivnog transporta vode (aldosteron, vazopresin, atriopeptid, progesteron, glukagon, kalcitonin, prostaglandini);
5) povećanje osmotskog pritiska tubularnog urina usled nereapsorbovanih osmotski aktivnih supstanci, kao što je glukoza (kontrinzularni hormoni);
6) promena protoka krvi kroz direktne sudove moždane supstance i samim tim akumulacija ili „ispiranje“ osmotski aktivnih supstanci iz intersticijuma (angiotenzin-II, kinini, prostaglandini, paratirin, vazopresin, atriopeptid).
Tubularna reapsorpcija elektrolita
tekstualna_polja
tekstualna_polja
arrow_upward
Tubularna reapsorpcija elektrolita, kao i vode, regulirana je pretežno hormonskim, a ne nervnim utjecajima.
Reapsorpcija natrijum u proksimalnim tubulima aktivira se aldosteronom i inhibira paratirin, u debelom dijelu uzlaznog ekstremiteta Henleove petlje reapsorpciju natriju aktiviraju vazopresin, glukagon, kalcitonin, a inhibiraju prostaglandini E. U distalnom dijelu tubuli, glavni regulatori transporta natrijuma su aldosteron (aktivacija), prostaglandini i atriopeptid (ugnjetavanje).
Regulacija cevnog transporta kalcijum,fosfat i djelimično magnezijum obezbjeđuju se uglavnom hormonima koji regulišu kalcijum. Paratirin ima nekoliko mjesta djelovanja u tubularnom aparatu bubrega. U proksimalnom tubulu (ravni dio), apsorpcija kalcija se odvija paralelno s transportom natrijuma i vode. Inhibicija reapsorpcije natrijuma u ovom dijelu pod utjecajem paratirina je praćena paralelnim smanjenjem reapsorpcije kalcija. Izvan proksimalnog tubula, paratirin selektivno pojačava reapsorpciju kalcija, posebno u distalnim zavijenim tubulima i kortikalnim sabirnim kanalima. Reapsorpciju kalcija također aktivira kalcitriol i inhibira kalcitonin. Apsorpciju fosfata u tubulima bubrega inhibiraju i paratirin (proksimalna reapsorpcija) i kalcitonin (distalna reapsorpcija), a pojačavaju je kalcitriol i somatotropin. Paratirin aktivira reapsorpciju magnezijuma u kortikalnom dijelu uzlaznog ekstremiteta Henleove petlje i inhibira proksimalnu reapsorpciju. bikarbonat.
Ljudski ekskretorni sistem vrši izlučivanje metaboličkih produkata u ljudsko tijelo. Rad organa ljudskog ekskretornog sistema ima svoje mehanizme za izlučivanje metaboličkih proizvoda nastalih u procesu evolucije, a to su filtracija, reapsorpcija i sekrecija.
ljudski ekskretorni sistem
Obavlja se izlučivanje metaboličkih produkata iz tijela, koji se sastoje od bubrega, uretera, mjehura i uretre.
Bubrezi se nalaze u retroperitonealnom prostoru u lumbalnoj regiji i imaju oblik graha.
Ovo je upareni organ koji se sastoji od kortikale i medule, zdjelice i prekriven je fibroznom membranom. Karlica bubrega sastoji se od male i velike čašice, a iz nje izlazi mokraćovod, koji doprema mokraću do mokraćne bešike i kroz uretru se konačni urin izlučuje iz organizma.
Bubrezi su uključeni u metaboličke procese, a njihova uloga u osiguravanju ravnoteže vode u tijelu, održavanju acido-bazne ravnoteže je osnovna za puno postojanje čovjeka.
Građa bubrega je veoma složena i njegov strukturni element je nefron.

Ima složenu strukturu i sastoji se od proksimalnog kanala, tijela nefrona, Henleove petlje, distalnog kanala i sabirnog kanala koji stvara mokraćovode. Reapsorpcija u bubrezima prolazi kroz tubule proksimalnog, distalnog dijela i Henleovu petlju.
Mehanizam reapsorpcije
Molekularni mehanizmi prolaska tvari u procesu reapsorpcije su:
- difuzija;
- endocitoza;
- pinocitoza;
- pasivni transport;
- aktivni transport.
Za reapsorpciju je od posebnog značaja aktivni i pasivni transport i pravac reapsorbovanih supstanci duž elektrohemijskog gradijenta i prisustvo nosača za supstance, rad ćelijskih pumpi i druge karakteristike.
Supstanca ide protiv elektrohemijskog gradijenta sa utroškom energije za njegovu implementaciju i kroz posebne transportne sisteme. Priroda kretanja je transcelularna, koja se odvija ukrštanjem apikalne i bazolateralne membrane. Takvi sistemi su:
- Primarni aktivni transport, koji se odvija uz pomoć energije razgradnje ATP-a. Koriste ga joni Na+, Ca+, K+, H+.
- Sekundarni aktivni transport odvija se zbog razlike u koncentraciji jona natrijuma u citoplazmi i u lumenu tubula, a ta razlika se objašnjava oslobađanjem natrijevih jona u međućelijsku tekućinu uz utrošak energije cijepanja ATP-a. Koriste ga aminokiseline, glukoza.
Prolazi duž gradijenata: elektrohemijskog, osmotskog, koncentracijskog, a njegova implementacija ne zahtijeva utrošak energije i stvaranje nosača. Supstance koje ga koriste su Cl-joni. Kretanje tvari je paraćelijsko. To je kretanje kroz ćelijsku membranu, koja se nalazi između dvije ćelije. Karakteristični molekularni mehanizmi su difuzija, transport sa rastvaračem.
Proces reapsorpcije proteina odvija se unutar ćelijske tekućine, a nakon cijepanja na aminokiseline, one ulaze u međućelijsku tekućinu, što nastaje kao rezultat pinocitoze.
Vrste reapsorpcije
Reapsorpcija je proces koji se odvija u tubulima. A tvari koje prolaze kroz tubule imaju različite nosače i mehanizme.

U bubrezima se dnevno formira od 150 do 170 litara primarnog urina, koji prolazi kroz proces reapsorpcije i vraća se u organizam. Tvari s visoko dispergiranim komponentama ne mogu proći kroz membranu tubula i u procesu reapsorpcije ući u krv s drugim tvarima.
proksimalna reapsorpcija
U proksimalnom nefronu, koji se nalazi u korteksu bubrega, odvija se reapsorpcija glukoze, natrijuma, vode, aminokiselina, vitamina i proteina.

Proksimalni tubul formiran je od epitelnih ćelija koje imaju apikalnu membranu i četkicu, a okrenut je prema lumenu bubrežnih tubula. Bazalna membrana formira nabore koji formiraju bazalni labirint, a kroz njih primarni urin ulazi u peritubularne kapilare. Ćelije su međusobno čvrsto povezane i formiraju prostor koji se proteže kroz međućelijski prostor tubula, a naziva se bazolateralni labirint.
Natrijum se reapsorbuje u složenom procesu u tri koraka i nosilac je drugih supstanci.
Reapsorpcija jona, glukoze i aminokiselina u proksimalnom tubulu
Glavni koraci u reapsorpciji natrijuma su:
- prolaz kroz apikalnu membranu. Ovo je faza pasivnog transporta natrijuma, kroz Na-kanale i Na-nosače. Joni natrija ulaze u ćeliju preko membranskih hidrofilnih proteina koji formiraju Na kanale.
- Ulazak ili prolazak kroz membranu povezan je sa izmjenom Na+ za vodonik, na primjer, ili sa njegovim ulaskom kao nosač glukoze, aminokiseline.
- prolaz kroz bazalnu membranu. Ovo je faza aktivnog transporta Na+, kroz Na+/K+ pumpe uz pomoć enzima ATP, koji, kada se razgradi, oslobađa energiju. Natrijum, koji se reapsorbuje u bubrežnim tubulima, stalno se vraća u metaboličke procese i njegova koncentracija u ćelijama proksimalnog tubula je niska.
Reapsorpcija glukoze se odvija kroz sekundarni aktivni transport, a njen unos je olakšan prenosom kroz Na-pumpu, te se potpuno vraća u metaboličke procese u organizmu. Povišena koncentracija glukoze se ne resorbuje u potpunosti u bubrezima i izlučuje se konačnim urinom.

Reapsorpcija aminokiselina odvija se slično kao i glukoza, ali složena organizacija aminokiselina zahtijeva učešće posebnih nosača za svaku aminokiselinu za manje od 5-7 dodatnih.
Reapsorpcija u Henleovoj petlji
Kroz Henleovu petlju prolazi i proces reapsorpcije u njenim uzlaznim i silaznim dijelovima za vodu i ione je različit.
Filtrat, ulazeći u silazni dio petlje, spuštajući se duž njega, oslobađa vodu zbog različitog gradijenta tlaka i zasićen je ionima natrija i klora. U ovom dijelu voda se reapsorbuje, te je nepropusna za jone. Uzlazni dio je nepropustan za vodu i pri prolasku kroz njega dolazi do razrjeđivanja primarnog urina, dok se u silaznom koncentriše.
Distalna reapsorpcija
Ovaj dio nefrona nalazi se u korteksu bubrega. Njegova funkcija je da reapsorbuje vodu koja se sakuplja u primarnom urinu i reapsorbuje ione natrija. Distalna reapsorpcija je razrjeđivanje primarnog urina i formiranje konačnog urina iz filtrata.
Ulazeći u distalni tubul, primarni urin u zapremini od 15% nakon reapsorpcije u bubrežnim tubulima iznosi 1% ukupne zapremine. Sakupljajući se nakon toga u sabirni kanal, razrjeđuje se i formira se konačni urin.
Neurohumoralna regulacija reapsorpcije
Reapsorpciju u bubrezima regulišu simpatički nervni sistem i tiroidni hormoni, hipotalamus-hipofiza i androgeni.
Reapsorpcija natrijuma, vode, glukoze se povećava uz ekscitaciju simpatičkog i vagusnog živca.
Distalni tubuli i sabirni kanalići reapsorbuju vodu u bubrezima pod uticajem antidiuretičkog hormona ili vazopresina, koji se sa smanjenjem vode u organizmu u velikim količinama povećava, a povećava se i propusnost zidova tubula.
Aldosteron povećava reapsorpciju kalcijuma, hlorida i vode, kao i atriopeptid, koji se proizvodi u desnoj pretkomori. Do inhibicije reapsorpcije natrijuma u proksimalnom nefronu dolazi kada uđe paratirin.
Aktivacija reapsorpcije natrijuma uzrokovana je hormonima:
- vazopresin.
- Glucogan.
- kalcitonin.
- Aldosteron.
Do inhibicije reapsorpcije natrijuma dolazi tokom proizvodnje hormona:
- Prostaglandin i prostaglandin E.
- Atriopeptid.
Moždana kora reguliše izlučivanje ili inhibiciju urina.
Tubularnu reapsorpciju vode provode mnogi hormoni odgovorni za propusnost membrana distalnog nefrona, regulaciju njegovog transporta kroz tubule i još mnogo toga.
Važnost reapsorpcije
Praktična primjena naučnih saznanja o tome šta je reapsorpcija u medicini omogućila je dobivanje informacijske potvrde o radu ekskretornog sistema tijela i sagledavanje njegovih unutrašnjih mehanizama. prolazi kroz veoma složene mehanizme i uticaj okoline, genetske abnormalnosti. I ne ostaju neprimijećeni kada se u njihovoj pozadini pojave problemi. Jednom rečju, zdravlje je veoma važno. Pratite njega i sve procese koji se odvijaju u tijelu.
Detalji
Reapsorpcija je transport tvari iz lumena bubrežnih tubula u krv teče kroz peritubularne kapilare. Reapsorbirano 65% primarnog volumena urina(oko 120 l/dan. Bilo je 170 l, izdvojeno je 1,5): voda, mineralne soli, sve potrebne organske komponente (glukoza, aminokiseline). Transport pasivno(osmoza, difuzija duž elektrohemijskog gradijenta) i aktivan(primarni aktivni i sekundarni aktivni uz učešće molekula nosača proteina). Transportni sistemi su isti kao u tankom crijevu.
Granične supstance - obično se potpuno reapsorbuju(glukoza, aminokiseline) i izlučuju se urinom samo ako njihova koncentracija u krvnoj plazmi premašuje graničnu vrijednost (tzv. "prag eliminacije"). Za glukozu, prag eliminacije je 10 mmol/l (pri normalnoj koncentraciji glukoze u krvi od 4,4-6,6 mmol/l).
Supstance bez praga - uvijek se izlučuju bez obzira na njihovu koncentraciju u krvnoj plazmi. Ne reapsorbuju se ili se samo djelimično reapsorbuju, kao što je urea i drugi metaboliti.
Mehanizam rada različitih dijelova bubrežnog filtera.
1. u proksimalnom tubulu nastaje proces koncentriranja glomerularnog filtrata, a najvažnija tačka je aktivna apsorpcija soli. Uz pomoć aktivnog transporta, oko 67% Na+ se reapsorbira iz ovog dijela tubula. Gotovo proporcionalna količina vode i nekih drugih otopljenih tvari, poput hloridnih jona, pasivno prati jone natrija. Dakle, prije nego što filtrat dođe do Henleove petlje, oko 75% tvari se reapsorbira iz njega. Kao rezultat, tubularna tekućina postaje izosmotična u odnosu na krvnu plazmu i tkivne tekućine.
Proksimalni tubul je idealan za intenzivna reapsorpcija soli i vode. Brojne mikrovile epitela čine takozvanu četkicu koja pokriva unutrašnju površinu lumena bubrežnog tubula. Ovakvim rasporedom upijajuće površine izuzetno se povećava površina stanične membrane i kao rezultat toga se olakšava difuzija soli i vode iz lumena tubula u epitelne stanice.
2. Silazni ud Henleove petlje i dio uzlaznog ekstremiteta nalazi u unutrašnjem sloju medula, sastoje se od vrlo tankih ćelija koje nemaju četkicu, a broj mitohondrija je mali. Morfologija tankih presjeka nefrona ukazuje na odsustvo aktivnog prijenosa otopljenih tvari kroz zid tubula. U ovom području nefrona NaCl vrlo slabo prodire kroz zid tubula, urea je nešto bolja, a voda prolazi bez poteškoća.
3. Zid tankog dijela uzlaznog ekstremiteta Henleove petlje također neaktivan u pogledu transporta soli. Ipak, ima visoku propusnost za Na+ i Cl-, ali je slabo propusna za ureu i skoro nepropusna za vodu.
4. Debeli dio uzlaznog ekstremiteta Henleove petlje, koji se nalazi u bubrežnoj meduli, razlikuje se od ostatka navedene petlje. Obavlja aktivan prijenos Na + i Cl - iz lumena petlje u intersticijski prostor. Ovaj dio nefrona, zajedno sa ostatkom uzlaznog koljena, izuzetno je slabo propustljiv za vodu. Zbog reapsorpcije NaCl, tečnost ulazi u distalni tubul donekle hipoosmotično u odnosu na tkivnu tečnost.
5. Kretanje vode kroz zid distalnog tubula- proces je složen. Distalni tubul je od posebnog značaja za transport K+, H+ i NH3 iz tkivne tečnosti u lumen nefrona i transport Na+, Cl- i H2O iz lumena nefrona u tkivnu tečnost. Pošto se soli aktivno „ispumpavaju“ iz lumena tubula, voda ih pasivno prati.
6. sabirni kanal propustljiv za vodu, omogućavajući mu da prođe iz razrijeđenog urina u koncentriraniju tkivnu tekućinu bubrežne medule. Ovo je završna faza u formiranju hiperosmotskog urina. Reapsorpcija NaCl se također javlja u kanalu, ali zbog aktivnog prijenosa Na+ kroz zid. Za soli, sabirni kanal je nepropustan, a za vodu njegova propusnost varira. Važna karakteristika distalnog dijela sabirnog kanala, koji se nalazi u unutrašnjoj meduli bubrega, je njegova visoka permeabilnost za ureu.

Mehanizam reapsorpcije glukoze.
Proksimalno(1/3) reapsorpcija glukoze se vrši uz pomoć specijalni nosači četkice apikalne membrane epitelnih ćelija. Ovi nosači prenose glukozu samo ako se vežu i transportuju natrijum. Pasivno kretanje natrijuma duž gradijenta koncentracije u ćelije dovodi do transporta preko membrane i nosača sa glukozom.
Za provedbu ovog procesa potrebna je niska koncentracija natrijuma u epitelnoj ćeliji, što stvara koncentracijski gradijent između vanjskog i unutarćelijskog okruženja, što se osigurava energetski ovisnim radom. pumpa sa bazalnom membranom natrijum-kalijum.
Ova vrsta transporta se zove sekundarno aktivno, ili symport, odnosno zajednički pasivni transport jedne supstance (glukoze) zbog aktivnog transporta druge (natrijuma) pomoću jednog nosača. Sa viškom glukoze u primarnom urinu, može doći do potpunog opterećenja svih molekula nosača i glukoza se više ne može apsorbirati u krv.
Ovu situaciju karakteriše maksimalni cevni transport materije» (Tm glukoza), što odražava maksimalno opterećenje tubularnih transportera pri određenoj koncentraciji tvari u primarnom urinu i, shodno tome, u krvi. Ova vrijednost se kreće od 303 mg/min kod žena do 375 mg/min kod muškaraca. Vrijednost maksimalnog tubularnog transporta odgovara konceptu "praga bubrežnog izlučivanja".
Prag bubrežne eliminacije nazovi to koncentracija supstance u krvi i, shodno tome, u primarnom urinu, pri čemu se više ne može potpuno reapsorbirati u tubulima i pojavljuje se u konačnom urinu. Takve tvari za koje se može naći prag eliminacije, odnosno potpuno se reapsorbiraju pri niskim koncentracijama u krvi, a ne u potpunosti u povišenim koncentracijama, nazivaju se pragom. Primjer je glukoza, koja se potpuno apsorbira iz primarnog urina pri koncentracijama u plazmi ispod 10 mmol/l, ali se pojavljuje u konačnom urinu, odnosno nije potpuno reapsorbirana, kada je njen sadržaj u krvnoj plazmi iznad 10 mmol/l. shodno tome, za glukozu, prag eliminacije je 10 mmol/l.

Mehanizmi izlučivanja u bubrežnom filteru.
Sekrecija je transport supstanci iz krvi teče kroz peritubularne kapilare u lumen bubrežnih tubula. Transport je pasivan i aktivan. Izlučuju se ioni H +, K +, amonijak, organske kiseline i baze (na primjer, strane tvari, posebno lijekovi: penicilin itd.). Sekrecija organskih kiselina i baza odvija se putem sekundarno aktivnog mehanizma ovisnog o natriju.

lučenje jona kalijuma.
Većina lako filtriranih jona kalija u glomerulu je obično reapsorbira iz filtrata u proksimalnim tubulima i Henleovim petljama. Brzina aktivne reapsorpcije u tubulu i petlji se ne smanjuje čak ni kada se koncentracija K+ u krvi i filtratu snažno poveća kao odgovor na prekomjernu potrošnju ovog jona u tijelu.
Međutim, distalni tubuli i sabirni kanalići su sposobni ne samo da reapsorbuju, već i da luče jone kalija. Lučenjem kalija, ove strukture teže postizanju homeostaze jona u slučaju da neuobičajeno velika količina ovog metala uđe u tijelo. Čini se da transport K+ zavisi od njegovog ulaska u tubularne ćelije iz tkivne tečnosti, zbog aktivnosti uobičajene Nar+-Ka+ pumpe, uz curenje K+ iz citoplazme u tubularni fluid. Kalijum može jednostavno da difunduje duž elektrohemijskog gradijenta iz ćelija bubrežnih tubula u lumen, jer je tubularna tečnost elektronegativna u odnosu na citoplazmu. Lučenje K+ kroz ove mehanizme stimuliše adrenokortikalni hormon aldosteron, koji se oslobađa kao odgovor na povećanje sadržaja K+ u krvnoj plazmi.
Tubularna reapsorpcija je proces reapsorpcije vode i tvari iz urina sadržanih u lumenu tubula u limfu i krv.
Većina molekula se reapsorbuje u proksimalnom nefronu. Ovdje se gotovo u potpunosti apsorbiraju aminokiseline, glukoza, vitamini, proteini, mikroelementi, značajna količina Na+, C1-, HCO3- jona i mnoge druge tvari.
Elektroliti i voda se apsorbiraju u Henleovoj petlji, distalnom tubulu i sabirnim kanalićima.
Aldosteron stimuliše reapsorpciju Na+ i izlučivanje K+ i H+ u bubrežne tubule u distalnom nefronu, u distalnom tubulu i kortikalnim sabirnim kanalima.
Vasopresin podstiče reapsorpciju vode iz distalnih uvijenih tubula i sabirnih kanala.
Uz pomoć pasivnog transporta, voda, hlor i urea se reapsorbuju.
Aktivni transport je prijenos tvari protiv elektrohemijskog i koncentracijskog gradijenta. Osim toga, razlikuju se primarno aktivni i sekundarno aktivni transport. Primarni aktivni transport nastaje trošenjem ćelijske energije. Primjer je prijenos Na+ jona pomoću enzima Na+/K+-ATPaze, koji koristi energiju ATP-a. U sekundarnom aktivnom transportu, prijenos tvari se vrši na račun transportne energije druge tvari. Glukoza i aminokiseline se reapsorbuju mehanizmom sekundarnog aktivnog transporta.
Vrijednost maksimalnog tubularnog transporta odgovara starom konceptu "praga bubrežnog izlučivanja". Za glukozu, ova vrijednost je 10 mmol/l.
Supstance čija reapsorpcija ne ovisi o njihovoj koncentraciji u krvnoj plazmi nazivaju se negranične. To uključuje tvari koje se ili uopće ne reapsorbiraju (inulin, manitol) ili se malo reapsorbiraju i izlučuju urinom srazmjerno njihovom nakupljanju u krvi (sulfati).
Normalno, mala količina proteina ulazi u filtrat i ponovo se apsorbuje. Proces reapsorpcije proteina provodi se uz pomoć pinocitoze. Po ulasku u ćeliju, protein se hidrolizira enzimima lizosoma i pretvara u aminokiseline. Ne podliježu svi proteini hidrolizi, neki od njih prolaze u krv nepromijenjeni. Ovaj proces je aktivan i zahtijeva energiju. Pojava proteina u urinu naziva se proteinurija. Proteinurija se može pojaviti i u fiziološkim uslovima, na primjer, nakon teškog mišićnog rada. U osnovi, proteinurija se javlja u patologiji nefritisa, nefropatija i multiplog mijeloma.
Urea igra važnu ulogu u mehanizmima koncentracije urina, jer se slobodno filtrira u glomerulima. U proksimalnom tubulu, dio uree se pasivno reapsorbuje gradijentom koncentracije koji nastaje zbog koncentracije urina. Ostatak uree dospijeva u sabirne kanale. U sabirnim kanalićima, pod uticajem ADH, voda se reapsorbuje i povećava se koncentracija uree. ADH povećava propusnost zida za ureu, te ona prelazi u medulu bubrega, stvarajući ovdje otprilike 50% osmotskog tlaka. Iz intersticija urea difundira duž gradijenta koncentracije u Henleovu petlju i ponovo ulazi u distalne tubule i sabirne kanale. Tako se odvija intrarenalna cirkulacija ureje. U slučaju vodene diureze, apsorpcija vode u distalnom nefronu prestaje, a izlučuje se više uree. Dakle, njegovo izlučivanje zavisi od diureze.
Reapsorpcija slabih kiselina i baza zavisi od toga da li su u jonizovanom ili nejonizovanom obliku. Slabe baze i kiseline u jonizovanom stanju se ne reapsorbuju i izlučuju se urinom. Stupanj jonizacije baza se povećava u kiseloj sredini, pa se one brže izlučuju kiselim urinom, slabe kiseline se, naprotiv, brže izlučuju alkalnom urinom. Ovo je od velike važnosti, jer su mnoge ljekovite tvari slabe baze ili slabe kiseline. Stoga je u slučaju trovanja acetilsalicilnom kiselinom ili fenobarbitalom (slabe kiseline) potrebno primijeniti alkalne otopine (NaHCO3) kako bi se ove kiseline prešle u jonizirano stanje, čime bi se olakšalo njihovo brzo eliminiranje iz organizma. Za brzo izlučivanje slabih baza potrebno je u krv uvesti kisele produkte za zakiseljavanje urina.
Voda se reapsorbuje u svim dijelovima nefrona pasivno zbog transporta osmotski aktivnih supstanci: glukoze, aminokiselina, proteina, natrijuma, kalija, kalcija, jona hlora. Sa smanjenjem reapsorpcije osmotski aktivnih supstanci, smanjuje se i reapsorpcija vode. Prisustvo glukoze u konačnom urinu dovodi do povećanja diureze (poliurije).
Natrijum je glavni ion odgovoran za pasivnu apsorpciju vode. Natrijum je, kao što je već pomenuto, takođe neophodan za transport glukoze i aminokiselina. Osim toga, igra važnu ulogu u stvaranju osmotski aktivnog okruženja u intersticiju bubrežne medule, čime se koncentriše urin.
Protok natrijuma iz primarnog urina kroz apikalnu membranu u ćeliju tubulnog epitela odvija se pasivno duž elektrohemijskog i koncentracijskog gradijenta. Izlučivanje natrijuma iz ćelije kroz bazolateralne membrane se odvija aktivno uz pomoć Na+/K+-ATPaze. Budući da se energija staničnog metabolizma troši na prijenos natrijuma, njegov transport je primarno aktivan. Transport natrijuma u ćeliju može se odvijati kroz različite mehanizme. Jedna od njih je zamjena Na + za H + (protivstrujni transport, ili antiport). U ovom slučaju, jon natrijuma se prenosi unutar ćelije, a ion vodonika se prenosi van. Drugi način prijenosa natrijuma u ćeliju provodi se uz sudjelovanje aminokiselina, glukoze. Ovo je takozvani kotransport, ili simport. Djelomično je reapsorpcija natrijuma povezana sa izlučivanjem kalija.
Srčani glikozidi (strofantin K, oubain) su u stanju da inhibiraju enzim Na + / K + -ATPazu, koji osigurava prijenos natrijuma iz stanice u krv i transport kalija iz krvi u ćeliju.
Od velikog značaja u mehanizmima reapsorpcije vode i jona natrijuma, kao i koncentraciji urina, je rad tzv. rotaciono-protivstrujnog multiplikatorskog sistema. Nakon prolaska kroz proksimalni segment tubula, izotonični filtrat u smanjenom volumenu ulazi u Henleovu petlju. U ovom dijelu intenzivna reapsorpcija natrijuma nije praćena reapsorpcijom vode, jer su zidovi ovog segmenta slabo propusni za vodu čak i pod utjecajem ADH. U tom smislu dolazi do razrjeđivanja urina u lumenu nefrona i koncentracije natrijuma u intersticijumu. Razrijeđen urin u distalnom tubulu gubi višak tekućine, postaje izotoničan s plazmom. Smanjena zapremina izotonične mokraće ulazi u sabirni sistem koji radi u meduli, čiji je visok osmotski pritisak u intersticijumu posledica povećane koncentracije natrijuma. U sabirnim kanalima, pod uticajem ADH, nastavlja se reapsorpcija vode u skladu sa gradijentom koncentracije. Vasa recta u meduli funkcioniraju kao protustrujni sudovi za razmjenu, uzimajući natrij na putu do papila i otpuštajući ga prije nego što se vraća u kortikalni sloj. U dubini medule na ovaj način se održava visok sadržaj natrijuma, koji osigurava resorpciju vode iz sabirnog sistema i koncentraciju urina.
Funkcije tubularnog aparata bubrega(koji uključuje proksimalni tubul, petlju nefrona, distalni tubul i sabirne kanale) su:
- reapsorpcija dijela organskih i neorganskih supstanci filtriranih u glomerulu;
- izlučivanje u lumen tubula tvari sadržanih u krvi ili formiranih u stanicama tubula,
- koncentracija urina.
Reapsorpcija - To je reverzna apsorpcija različitih tvari iz lumena tubula u plazmu peritubularnih kapilara. Reapsorpcija se javlja u svim dijelovima tubula nefrona, u sabirnom kanalu i određena je strukturnim karakteristikama tubularnog epitela bubrega. Površina ćelija proksimalnog zavijenog tubula, okrenuta prema njegovom lumenu, ima gustu četkicu prekrivenu glikokaliksom, što povećava kontaktnu površinu membrane s tubularnom tekućinom za 40 puta. Postoje propusni čvrsti spojevi između ćelija ispod ivice četke.
Apikalni dio plazmaleme naziva se i luminalnim, ima visoku ionsku permeabilnost, sadrži različite proteine nosače i obezbjeđuje pretežno pasivan transport različitih supstanci.
Bazolateralni dio ćelije povećana zbog savijanja membrane i sadrži veliki broj mitohondrija, što određuje koncentraciju aktivnih transportnih sistema (jonskih pumpi) u njoj.
prag reapsorpcije odražava ovisnost apsorpcije tvari o njezinoj koncentraciji u krvnoj plazmi. Ako koncentracija tvari u plazmi ne prelazi određeni prag, tada će se ova tvar potpuno reapsorbirati u tubulima nefrona, ako je veća, onda se ne resorbira u potpunosti i pojavljuje se u konačnom urinu, što je povezano uz maksimalno zasićenje nosilaca.
primarni urin, prolazeći kroz tubule i cijevi za prikupljanje, prije nego što se pretvori u konačni urin, doživljava značajne promjene. Razlika nije samo u količini (od 180 litara ostaje 1-1,5 litara), već i u kvaliteti. Neke tvari potrebne tijelu potpuno nestaju iz mokraće ili postaju znatno manje. ide proces reapsorpcije. Koncentracija drugih supstanci se višestruko povećava: one se koncentrišu kada se voda ponovo apsorbira. Još druge supstance koje uopće nisu bile u primarnom urinu,
pojaviti na kraju. To se događa kao rezultat njihovog lučenja.
procesi reapsorpcije mogu biti aktivni ili pasivni. Za implementaciju aktivni proces neophodno je da postoje specifični transportni sistemi i energija. Pasivni procesi nastaju, po pravilu, bez trošenja energije prema zakonima fizike i hemije.
tubularna reapsorpcija javlja se u svim odjelima, ali njegov mehanizam u različitim dijelovima nije isti. Uslovno je moguće razlikovati C odjeljenja: proksimalni uvijeni tubul, petlja nefrona i distalni uvijeni tubul Sa zbirnom cijevi.
U proksimalnim izvijenim tubulima aminokiseline, glukoza, vitamini, proteini, elementi u tragovima se potpuno reapsorbuju. U istom delu se reapsorbuje oko 2/3 vode i neorganskih soli Na +, K + Ca2 +, Mg2 +, Cl-, HC07, tj. tvari koje su tijelu potrebne za svoje aktivnosti. Mehanizam reapsorpcije uglavnom je direktno ili indirektno povezan sa reapsorpcijom Na+.
Reapsorpcija natrijuma .
Većina Na+ se reapsorbuje protiv gradijenta koncentracije zbog energije ATP-a. Izvodi se reapsorpcija Na + u 3 faze: prijenos jona kroz apikalnu membranu tubularnih epitelnih stanica, transport do bazalnih ili lateralnih membrana i prijenos kroz ove membrane u međućelijsku tekućinu i u krv. Glavna pokretačka sila reapsorpcije je prijenos Na+ preko Na+,K+-ATPaze preko bazolateralne membrane. Ovo osigurava stalan odliv jona. Kao rezultat toga, Na + duž gradijenta koncentracije uz pomoć posebnih formacija endoplazmatskog retikuluma ulazi u membrane vraćene u međućelijsku sredinu. Kao rezultat ovog neprekidnog transportera, koncentracija iona unutar ćelije, a posebno u blizini apikalne membrane, postaje mnogo niža nego na njenoj drugoj strani, što doprinosi pasivnom ulasku Na+ u ćeliju duž ionskog gradijenta. Na ovaj način,
2 faze reapsorpcije natrijuma od strane tubularnih ćelija su pasivne i samo jedna, zadnja, zahtijeva energiju. Osim toga, dio Na+ se pasivno reapsorbuje duž međućelijskih prostora zajedno s vodom.
Glukoza. Glukoza se reapsorbuje zajedno sa transportom Na+. U apikalnoj membrani ćelija postoje posebne transporteri. Riječ je o proteinima molekulske težine 320.000, koji u početnim dijelovima proksimalnog tubula nose svaki Na+ i jedan molekul glukoze (postupno smanjenje koncentracije glukoze u urinu dovodi do toga da se u sljedećem području tubul dva Na+ se već koriste za prijenos jednog molekula glukoze). Ovaj proces je takođe vođen elektrohemijski gradijent Na+. Na suprotnoj strani ćelije, kompleks Na-glukoza-nosač se raspada na tri elementa. Kao rezultat toga, oslobođeni nosač se vraća na svoje prvobitno mjesto i ponovo stječe sposobnost da nosi nove komplekse Na+ i glukoze. U ćeliji se povećava koncentracija glukoze, zbog čega se formira koncentracijski gradijent koji je usmjerava na bazalno-lateralne membrane stanice i osigurava izlaz u međućelijsku tekućinu. Odavde glukoza ulazi u krvne kapilare i vraća se u opću cirkulaciju. Apikalna membrana sprječava da glukoza prođe natrag u lumen tubula. Transporteri glukoze nalaze se samo u proksimalnom tubulu, pa se glukoza reapsorbuje samo ovdje.
U redu na uobičajenom nivou glukoze u krvi, a time i njenoj koncentraciji u primarnom urinu, sva glukoza se reapsorbuje. Međutim, s povećanjem nivoa glukoze u krvi za više od 10 mmol/l (oko 1,8 g/l), kapacitet transportnih sistema postaje nedovoljan za reapsorpciju. Prvi tragovi nereapsorbirane glukoze u konačnom urinu se otkrivaju kada je njegova koncentracija u krvi prekoračena. Što je veća koncentracija glukoze u krvi, veća je i količina nereapsorbirane glukoze. Do njegove koncentracije od 3,5 g/l ovo povećanje još nije direktno proporcionalno, budući da neki od transportera još nisu uključeni u proces. ali, počevši od 3,5 g/l, izlučivanje glukoze u urinu postaje direktno proporcionalno njenoj koncentraciji u krvi. Kod muškaraca puno opterećenje reapsorpcionog sistema se opaža pri unosu 2,08 mmol/min (375 mg/min) glukoze, i kod žena- 1,68 mmol/min (303 mg/min) na osnovu 1,73 m2 telesne površine.
Amino kiseline. Reapsorpcija aminokiselina odvija se po istom mehanizmu kao i reapsorpcija glukoze. Potpuna reapsorpcija aminokiselina događa se već u početnim dijelovima proksimalnih tubula. Ovaj proces je povezan sa aktivnom reapsorpcijom Na+ kroz apikalnu membranu ćelija. Otkriveno 4 vrste transportnih sistema: a) za bazne b) za kiselinu c) za hidrofilne d) za hidrofobne aminokiseline. Iz ćelije aminokiseline prolaze pasivno duž gradijenta koncentracije kroz bazalnu membranu u međućelijsku tečnost, a odatle u krv. Pojava aminokiselina u urinu može biti posljedica kršenja transportnih sistema ili vrlo visoke koncentracije u krvi. U potonjem slučaju može doći do efekta koji po mehanizmu podsjeća na glukozuriju - preopterećenje transportnih sistema. Ponekad postoji konkurencija između kiselina istog tipa za zajednički nosač.
Vjeverice. Mehanizam reapsorpcije proteina značajno se razlikuje od mehanizma reapsorpcije opisanih spojeva. Jednom u primarnom urinu, mala količina proteina se normalno skoro potpuno reapsorbuje pinocitozom. U citoplazmi stanica proksimalnih tubula, proteini se razgrađuju uz sudjelovanje lizosomalnih enzima. Aminokiseline koje se formiraju prate gradijent koncentracije od ćelije do međućelijske tečnosti, a odatle do krvnih kapilara. Na taj način se do 30 mg proteina može reapsorbirati za 1 min. Ako su glomeruli oštećeni, više proteina ulazi u filtrat, a dio može ući u urin ( proteinurija).
tubularna sekrecija. U savremenoj fiziološkoj literaturi koja se tiče aktivnosti bubrega, termin sekrecija ima dva značenja. Prvo od njih opisuje proces prijenosa tvari kroz stanice iz krvi u lumen tubula u nepromijenjenom obliku, što povećava brzinu izlučivanja tvari putem bubrega. Sekunda- oslobađanje iz ćelije u krv ili u lumen tubula fiziološki aktivnih supstanci sintetiziranih u bubrezima (na primjer prostaglandini, bradikinin itd.) ili izlučenih tvari (na primjer, hipurinska kiselina).
Lučenje organskih i neorganskih supstanci- jedan od važnih procesa koji osiguravaju proces mokrenja. Kod riba nekih vrsta, glomeruli su odsutni u bubrezima. U takvim slučajevima sekret igra vodeću ulogu u radu bubrega. U bubrezima većine drugih klasa kralježnjaka, uključujući sisare, sekrecija osigurava oslobađanje dodatnih količina određenih tvari iz krvi u lumen tubula, koje se mogu filtrirati u bubrežnim glomerulima.
Na ovaj način, sekrecija se ubrzava izlučivanje preko bubrega nekih stranih supstanci, krajnjih produkata metabolizma, jona. Organske kiseline (penicilin, paraaminohipurna kiselina - PAG, diodrast, mokraćna kiselina), organske baze (holin, gvanidin) i neorganske supstance (kalijum) luče se u bubrezima sisara. Bubrezi glomerularnih i aglomerularnih morskih koštanih riba sposobni su da luče ione magnezijuma, kalcijuma i sulfata. Mjesta lučenja različitih tvari se razlikuju. U bubrezima svih kralježnjaka stanice proksimalnog segmenta nefrona, posebno njegovog direktnog dijela, služe kao mjesto lučenja organskih kiselina i baza; lučenje kalija se događa uglavnom u stanicama distalnog uvijenog tubula i sabirnih kanala.
Mehanizam procesa lučenja organskih kiselina. Razmotrimo ovaj proces na primjeru izlučivanja PAH-a putem bubrega. Nakon unošenja PAH-a u krv, povećava se njegovo lučenje u bubrezima i pročišćavanje krvi iz njega značajno premašuje količinu pročišćavanja krvi od istovremeno primijenjenog inulina. To znači da se PAH ne samo filtrira u glomerulima, već, osim u glomerule, značajne količine ulaze u lumen nefrona. Eksperimentalno je pokazano da je ovaj proces posljedica izlučivanja PAH-a iz krvi u lumen proksimalnih tubula. U ćelijskoj membrani ovog tubula, okrenutom prema međućelijskoj tekućini, nalazi se nosač ( kotransporter), sa visokim afinitetom za PAG. U prisustvu PAG-a nastaje kompleks nosača sa PAG-om, koji se kreće u membrani i razgrađuje na njenoj unutrašnjoj površini, oslobađajući PAG u citoplazmu, a nosač ponovo dobija sposobnost da se kreće na vanjsku površinu membrane. i kombinirati s novom PAG molekulom. Mehanizam sekrecije organske kiseline uključuje niz koraka. Bazalna plazma membrana sadrži Na+, K+-ATPazu, koja uklanja Na+ ione iz ćelije i podstiče ulazak K+ jona u ćeliju. Niža koncentracija Na+ jona u citoplazmi omogućava ionima Na+ da uđu u ćeliju duž gradijenta koncentracije uz učešće kotransportera natrijuma. Jedan od tipova takvog kotransportera pospješuje ulazak α-ketoglutarata i Na+ kroz bazalnu plazma membranu. Ista membrana ima anjonski izmjenjivač koji uklanja α-ketoglutarat iz citoplazme u zamjenu za paraaminohipurat (PAG), diodrast ili neke druge organske kiseline koje iz intersticijske tekućine dolaze u ćeliju. Ova supstanca se kreće kroz ćeliju prema luminalnoj membrani i kroz nju prolazi u lumen tubula mehanizmom olakšane difuzije.
Respiratorna depresija cijanidi, razdvajanje disanja i oksidativna fosforilacija dinitrofenolom smanjuje i zaustavlja sekreciju. U normalnim fiziološkim uslovima, nivo sekrecije zavisi od broja nosača u membrani. Lučenje PAG-a se povećava proporcionalno povećanju koncentracije PAG-a u krvi sve dok svi molekuli nosači ne budu zasićeni PAG-om. Maksimalna brzina transporta PAG-a se postiže kada je količina PAG-a dostupna za transport jednaka broju molekula nosača koji mogu formirati kompleks sa PAG-om. Ova vrijednost je definirana kao maksimalna sposobnost transporta PAG - Ttrans. PAH koji ulazi u ćeliju kreće se duž citoplazme do apikalne membrane i kroz nju se posebnim mehanizmom oslobađa u lumen tubula.
Ulaznica 15
Prethodna3456789101112131415161718Sljedeća
BUBREZI I NJIHOVE FUNKCIJE
tubularna reapsorpcija
Početni stadijum mokrenja, koji dovodi do filtracije svih niskomolekularnih komponenti krvne plazme, neminovno se mora kombinovati sa postojanjem sistema u bubregu koji reapsorbuju sve supstance vredne za organizam. U normalnim uslovima, u ljudskom bubregu dnevno se formira do 180 litara filtrata, a izluči se 1,0-1,5 litara urina, ostatak tečnosti se apsorbuje u tubulima. Uloga ćelija različitih segmenata nefrona u reapsorpciji nije ista. Eksperimenti sprovedeni na životinjama sa ekstrakcijom tečnosti iz različitih delova nefrona mikropipetom omogućili su da se razjasne karakteristike reapsorpcije različitih supstanci u različitim delovima bubrežnih tubula (slika 12.6). U proksimalnom segmentu nefrona skoro potpuno se reapsorbuju aminokiseline, glukoza, vitamini, proteini, elementi u tragovima, značajna količina jona Na+, CI-, HCO3. U kasnijim slučajevima nefrona, elektroliti i voda se pretežno apsorbuju.
Reapsorpcija natrijuma i hlora je najznačajniji proces u smislu zapremine i potrošnje energije. U proksimalnom tubulu, kao rezultat reapsorpcije većine filtriranih supstanci i vode, smanjuje se volumen primarnog urina, a oko 1/3 tekućine filtrirane u glomerulima ulazi u početni dio nefronske petlje. Od ukupne količine natrijuma koja je ušla u nefron tokom filtracije, do 25% se apsorbira u nefronskoj petlji, oko 9% u distalnom izvijenom tubulu, a manje od 1% se reapsorbira u sabirnim kanalima ili izluči urinom.
Reapsorpciju u distalnom segmentu karakteriše činjenica da ćelije nose manju količinu jona nego u proksimalnom tubulu, ali nasuprot većem gradijentu koncentracije. Ovaj segment nefrona i sabirni kanali igraju ključnu ulogu u regulaciji volumena izlučenog urina i koncentracije osmotski aktivnih supstanci u njemu (osmotska koncentracija1). U konačnom urinu koncentracija natrijuma može se smanjiti na 1 mmol/l u poređenju sa 140 mmol/l u krvnoj plazmi. U distalnom tubulu, kalijum se ne samo reapsorbuje, već se i luči kada ga ima u organizmu u višku.
U proksimalnom nefronu dolazi do reapsorpcije natrijuma, kalija, hlora i drugih supstanci kroz membranu zida tubula, koja je visoko propusna za vodu. Nasuprot tome, u debeloj uzlaznoj nefronskoj petlji, distalnim uvijenim tubulima i sabirnim kanalima, dolazi do reapsorpcije jona i vode kroz zid tubula, koji je manje propusn za vodu; permeabilnost membrane za vodu u pojedinim dijelovima nefrona i sabirnih kanala može se regulisati, a vrijednost propusnosti varira u zavisnosti od funkcionalnog stanja organizma (fakultativna reapsorpcija). Pod uticajem impulsa koji dolaze kroz eferentne nerve, i pod dejstvom biološki aktivnih supstanci, reguliše se reapsorpcija natrijuma i hlora u proksimalnom nefronu. To je posebno izraženo u slučaju povećanja volumena krvi i ekstracelularne tekućine, kada smanjenje reapsorpcije u proksimalnom tubulu doprinosi povećanju izlučivanja iona i vode, a samim tim i obnavljanju vode-soli. balans. U proksimalnom tubulu izoosmija je uvijek očuvana. Zid tubula je propustljiv za vodu, a volumen reapsorbirane vode određen je količinom reapsorbiranih osmotski aktivnih tvari, iza kojih se voda kreće duž osmotskog gradijenta. U krajnjim dijelovima distalnog segmenta nefrona i sabirnim kanalićima vazopresin reguliše propusnost zida tubula za vodu.
Fakultativna reapsorpcija vode zavisi od osmotske permeabilnosti zida tubula, veličine osmotskog gradijenta i brzine kretanja tečnosti kroz tubul.
Za karakterizaciju apsorpcije različitih supstanci u bubrežnim tubulima neophodna je ideja o pragu izlučivanja.
Tvari bez praga oslobađaju se u bilo kojoj koncentraciji u krvnoj plazmi (i, prema tome, u ultrafiltratu). Takve supstance su inulin, manitol. Prag za izlučivanje gotovo svih fiziološki važnih, vrijednih supstanci za organizam je različit. Dakle, oslobađanje glukoze u urinu (glukozurija) nastaje kada njena koncentracija u glomerularnom filtratu (i u krvnoj plazmi) prelazi 10 mmol / l. Fiziološko značenje ovog fenomena će biti otkriveno u opisu mehanizma reapsorpcije.
Mehanizmi tubularne reapsorpcije. Reverzna apsorpcija različitih tvari u tubulima osigurava se aktivnim i pasivnim transportom. Ako se supstanca reapsorbuje u odnosu na elektrohemijske i koncentracijske gradijente, proces se naziva aktivni transport. Postoje dvije vrste aktivnog transporta - primarno aktivno i sekundarno aktivno. Primarni aktivni transport naziva se kada se supstanca prenosi protiv elektrohemijskog gradijenta zbog energije ćelijskog metabolizma. Primjer je transport Na + jona, koji se odvija uz učešće Na + enzima, K + -ATPaze, koja koristi energiju ATP-a. Sekundarno-aktivan je prijenos tvari protiv gradijenta koncentracije, ali bez trošenja ćelijske energije direktno na ovaj proces; pa se glukoza, aminokiseline reapsorbuju. Iz lumena tubula ove organske tvari ulaze u stanice proksimalnog tubula uz pomoć posebnog nosača, koji nužno mora vezati ion Na+. Ovaj kompleks (nosač + organska tvar + Na+) potiče kretanje supstance kroz membranu četkice i njen ulazak u ćeliju. Pokretačka snaga za prijenos ovih supstanci preko apikalne plazma membrane je niža koncentracija natrijuma u citoplazmi ćelije u odnosu na lumen tubula. Gradijent koncentracije natrijuma nastaje zbog kontinuiranog aktivnog izlučivanja natrijuma iz ćelije u ekstracelularnu tečnost uz pomoć Na+, K+-ATPaze lokalizovane u bočnim i bazalnim membranama ćelije.
Reapsorpcija vode, klora i nekih drugih jona, uree se vrši pasivnim transportom - duž elektrohemijskog, koncentracijskog ili osmotskog gradijenta. Primjer pasivnog transporta je reapsorpcija hlora u distalnom zavijenom tubulu duž elektrohemijskog gradijenta stvorenog aktivnim transportom natrijuma. Voda se transportuje duž osmotskog gradijenta, a brzina njene apsorpcije zavisi od osmotske permeabilnosti zida tubula i razlike u koncentraciji osmotski aktivnih supstanci sa obe strane njegovog zida. U sadržaju proksimalnog tubula, zbog apsorpcije vode i tvari otopljenih u njoj, povećava se koncentracija uree, čija se mala količina reapsorbira u krv duž gradijenta koncentracije.
Dostignuća u oblasti molekularne biologije omogućila su da se uspostavi struktura molekula jonskih i vodenih kanala (akvaporina) receptora, autakoida i hormona, te se tako prodre u suštinu nekih ćelijskih mehanizama koji osiguravaju transport supstanci. kroz zid tubula. Osobine ćelija različitih delova nefrona su različite, svojstva citoplazmatske membrane u istoj ćeliji nisu ista. Apikalna membrana ćelije, okrenuta ka lumenu tubula, ima drugačije karakteristike od bazalne i bočne membrane, oprane međućelijskom tečnošću i u kontaktu sa krvnom kapilarom. Kao rezultat toga, apikalna i bazalna plazma membrana učestvuju u transportu supstanci na različite načine; specifično je i djelovanje biološki aktivnih tvari na obje membrane.
Razmotrite ćelijski mehanizam reapsorpcije jona koristeći Na+ kao primjer. U proksimalnom tubulu nefrona, apsorpcija Na+ u krv nastaje kao rezultat niza procesa, od kojih je jedan aktivni transport Na+ iz lumena tubula, a drugi pasivna reapsorpcija Na+ nakon i bikarbonat i C1- jone koji se aktivno transportuju u krv. Uvođenjem jedne mikroelektrode u lumen tubula, a druge - u peritubularnu tekućinu, utvrđeno je da se razlika potencijala između vanjske i unutrašnje površine zida proksimalnog tubula pokazala vrlo malom - oko 1,3 mV, u regionu distalnog tubula može dostići - 60 mV (sl. 12.7). Lumen oba tubula je elektronegativan, a u krvi (dakle, u ekstracelularnoj tečnosti) koncentracija Na+ je veća nego u tečnosti u lumenu ovih tubula, pa se reapsorpcija Na+ odvija aktivno protiv gradijenta. elektrohemijskog potencijala. Istovremeno, iz lumena tubula, Na + ulazi u ćeliju kroz natrijev kanal ili uz sudjelovanje nosača. Unutrašnji dio ćelije je negativno nabijen, a pozitivno nabijeni Na+ ulazi u ćeliju duž gradijenta potencijala, kreće se prema bazalnoj plazma membrani, kroz koju se natrijum pumpom izbacuje u međućelijsku tekućinu; gradijent potencijala preko ove membrane dostiže 70–90 mV.
Postoje supstance koje mogu uticati na pojedine elemente sistema reapsorpcije Na+. Tako je natrijum kanal u ćelijskoj membrani distalnog tubula i sabirnog kanala blokiran amiloridom i triamterenom, zbog čega Na+ ne može ući u kanal. Postoji nekoliko vrsta jonskih pumpi u ćelijama.
Tubularna reapsorpcija i njena regulacija
Jedna od njih je Na+, K+-ATPaza. Ovaj enzim se nalazi u bazalnoj i lateralnoj membrani ćelije i obezbeđuje transport Na+ iz ćelije u krv i ulazak K+ iz krvi u ćeliju. Enzim inhibiraju srčani glikozidi, kao što su strofantin, ouabain. U reapsorpciji bikarbonata važnu ulogu ima enzim karboanhidraza čiji je inhibitor acetazolamid – zaustavlja reapsorpciju bikarbonata koji se izlučuje urinom.
Filtriranu glukozu gotovo u potpunosti reapsorbiraju stanice proksimalnog tubula, a normalno se mala količina (ne više od 130 mg) izlučuje urinom dnevno. Proces reapsorpcije glukoze odvija se protiv visokog gradijenta koncentracije i sekundarno je aktivan. U apikalnoj (luminalnoj) ćelijskoj membrani glukoza se spaja s nosačem, koji također mora vezati Na+, nakon čega se kompleks transportuje kroz apikalnu membranu, odnosno glukoza i Na+ ulaze u citoplazmu. Apikalna membrana je visoko selektivna i jednosmjerno propusna i ne pušta glukozu ili Na+ natrag iz ćelije u lumen tubula. Ove supstance se kreću prema bazi ćelije duž gradijenta koncentracije. Prenos glukoze iz ćelije u krv kroz bazalnu plazma membranu ima karakter olakšane difuzije, a Na+ se, kao što je već navedeno, uklanja natrijum pumpom koja se nalazi u ovoj membrani.
Aminokiseline se gotovo u potpunosti reapsorbiraju u stanicama proksimalnog tubula. Postoje najmanje 4 sistema za transport aminokiselina iz lumena tubula u krv, reapsorbujući neutralne, dvobazne, dikarboksilne aminokiseline i aminokiseline. Svaki od ovih sistema osigurava apsorpciju određenog broja aminokiselina iste grupe. Dakle, sistem reapsorpcije dvobaznih aminokiselina je uključen u apsorpciju lizina, arginina, ornitina i eventualno cistina. Unošenjem viška jedne od ovih aminokiselina u krv počinje pojačano izlučivanje aminokiselina samo ove grupe putem bubrega. Transportni sistemi odvojenih grupa aminokiselina kontrolišu se odvojenim genetskim mehanizmima. Opisane su nasljedne bolesti čija je jedna od manifestacija pojačano izlučivanje određenih grupa aminokiselina (aminoacidurija).
Izlučivanje slabih kiselina i baza u urinu ovisi o njihovoj glomerularnoj filtraciji, reapsorpciji ili procesu sekrecije. Proces izlučivanja ovih supstanci u velikoj mjeri je određen "nejonskom difuzijom", čiji je utjecaj posebno izražen u distalnim tubulima i sabirnim kanalićima. Slabe kiseline i baze mogu postojati u zavisnosti od pH sredine u dva oblika - nejonizovanom i jonizovanom. Stanične membrane su propusnije za nejonizirane tvari. Mnoge slabe kiseline se brže izlučuju u alkalnom urinu, dok se slabe baze, naprotiv, izlučuju u kiselom urinu. Stepen jonizacije baza se povećava u kiseloj sredini, ali se smanjuje u alkalnoj. U nejoniziranom stanju ove tvari prodiru kroz membranske lipide u stanice, a zatim u krvnu plazmu, odnosno reapsorbuju se. Ako se pH vrijednost tubularne tekućine pomakne na kiselu stranu, tada se baze ioniziraju, slabo se apsorbiraju i izlučuju urinom. Nikotin je slaba baza, pri pH 8,1 50% se jonizuje, izlučuje se 3-4 puta brže kiselim (pH oko 5) nego alkalnim (pH 7,8) urinom. Proces "nejonske difuzije" utiče na izlučivanje slabih baza i kiselina, barbiturata i drugih lijekova putem bubrega.
Mala količina proteina filtriranog u glomerulima se reapsorbuje od strane ćelija proksimalnih tubula. Izlučivanje proteina u urinu normalno nije više od 20-75 mg dnevno, a u slučaju bolesti bubrega može se povećati i do 50 g dnevno. Povećanje izlučivanja proteina u urinu (proteinurija) može biti posljedica kršenja njihove reapsorpcije ili povećanja filtracije.
Za razliku od reapsorpcije elektrolita, glukoze i aminokiselina, koji, nakon prodiranja kroz apikalnu membranu, dospiju u bazalnu plazma membranu nepromijenjeni i transportiraju se u krv, reapsorpcija proteina je osigurana fundamentalno drugačijim mehanizmom. Protein ulazi u ćeliju pinocitozom. Filtrirani proteinski molekuli se adsorbiraju na površini apikalne ćelijske membrane, dok membrana učestvuje u formiranju pinocitne vakuole. Ova vakuola se kreće prema bazalnom dijelu ćelije. U perinuklearnoj regiji, gdje je lokaliziran lamelarni kompleks (Golgijev aparat), vakuole se mogu spojiti s lizosomima, koji imaju visoku aktivnost niza enzima. U lizosomima se zarobljeni proteini cijepaju, a nastale aminokiseline, dipeptidi se uklanjaju u krv kroz bazalnu plazma membranu. Međutim, treba naglasiti da se ne hidrolizuju svi proteini tokom transporta, a neki od njih se prenose u krv u nepromijenjenom obliku.
Određivanje količine reapsorpcije u tubulima bubrega. Reapsorpcija supstanci, odnosno njihov transport (T) iz lumena tubula u tkivnu (međućelijsku) tečnost i u krv, tokom reapsorpcije R (TRX) određuje se razlikom između količine supstance X. (F∙Px∙fx) filtrirana u glomerule, a količina supstance izlučena u urinu (UX ∙V).
TRX =F∙px.fx ─Ux∙V,
gdje je F volumen glomerularne filtracije, fx je udio supstance X koja nije vezana za proteine u plazmi u odnosu na njenu ukupnu koncentraciju u krvnoj plazmi, P je koncentracija supstance u krvnoj plazmi, U je koncentracija supstance u urin.
Prema gornjoj formuli izračunava se apsolutna količina reapsorbirane supstance. Prilikom izračunavanja relativne reapsorpcije (% R) određuje se udio tvari koja je podvrgnuta reapsorpciji u odnosu na količinu tvari filtrirane u glomerulima:
% R= (1 - EFX)∙100.
Za procjenu kapaciteta reapsorpcije proksimalnih tubularnih stanica važno je odrediti maksimalnu vrijednost transporta glukoze (TmG). Ova vrijednost se mjeri kada je sistem njegovog tubularnog transporta potpuno zasićen glukozom (vidi sliku 12.5). Da bi se to učinilo, otopina glukoze se ulijeva u krv i time se povećava njena koncentracija u glomerularnom filtratu sve dok se značajna količina glukoze ne počne izlučivati urinom:
TmG=F∙PG-UG∙V,
gdje je F glomerularna filtracija, PG je koncentracija glukoze u krvnoj plazmi, a UG je koncentracija glukoze u urinu; Tm - maksimalni cevni transport ispitivane supstance. Vrijednost TmG karakterizira puno opterećenje sistema za transport glukoze; kod muškaraca ova vrijednost iznosi 375 mg/min, a kod žena - 303 mg/min, na osnovu 1,73 m2 tjelesne površine.
tubularna reapsorpcija
Primarni urin se pretvara u konačni urin kroz procese koji se odvijaju u bubrežnim tubulima i sabirnim bačvama. U ljudskom bubregu dnevno se formira 150-180 litara filma, odnosno primarnog urina, a izluči se 1,0-1,5 litara urina. Ostatak tečnosti se apsorbuje u tubulima i sabirnim kanalima.
Tubularna reapsorpcija je proces reapsorpcije vode i tvari iz urina sadržanih u lumenu tubula u limfu i krv. Glavna svrha reapsorpcije je da se u tijelu održe sve vitalne tvari u potrebnim količinama. Reapsorpcija se javlja u svim dijelovima nefrona. Većina molekula se reapsorbuje u proksimalnom nefronu. Ovdje se gotovo u potpunosti apsorbiraju aminokiseline, glukoza, vitamini, proteini, mikroelementi, značajna količina Na+, C1-, HCO3- jona i mnoge druge tvari.
Shema tubularne reapsorpcije
Elektroliti i voda se apsorbiraju u Henleovoj petlji, distalnom tubulu i sabirnim kanalićima. Ranije se smatralo da je reapsorpcija u proksimalnom tubulu obavezna i neregulisana. Sada je dokazano da ga regulišu i nervni i humoralni faktori.
Reapsorpcija različitih supstanci u tubulima može se odvijati pasivno i aktivno. Pasivni transport se odvija bez potrošnje energije duž elektrohemijskog, koncentracijskog ili osmotskog gradijenta. Uz pomoć pasivnog transporta, voda, hlor i urea se reapsorbuju.
Aktivni transport je prijenos tvari protiv elektrohemijskog i koncentracijskog gradijenta. Osim toga, razlikuju se primarno aktivni i sekundarno aktivni transport. Primarni aktivni transport nastaje trošenjem ćelijske energije. Primjer je prijenos Na + jona uz pomoć enzima Na +, K + - ATPaze, koji koristi energiju ATP-a. U sekundarnom aktivnom transportu, prijenos tvari se vrši na račun transportne energije druge tvari. Glukoza i aminokiseline se reapsorbuju mehanizmom sekundarnog aktivnog transporta.
Glukoza. Dolazi iz lumena tubula u ćelije proksimalnog tubula uz pomoć posebnog nosača, koji nužno mora pričvrstiti ion Ma4'. Kretanje ovog kompleksa u ćeliju odvija se pasivno duž elektrohemijskog i koncentracijskog gradijenta za jone Na+. Niska koncentracija natrijuma u ćeliji, koja stvara gradijent njene koncentracije između vanjskog i intracelularnog okruženja, osigurava se radom natrijum-kalijum pumpe bazalne membrane.
U ćeliji se ovaj kompleks raspada na sastavne komponente. Unutar bubrežnog epitela stvara se visoka koncentracija glukoze, pa u budućnosti, duž gradijenta koncentracije, glukoza prelazi u intersticijsko tkivo. Ovaj proces se izvodi uz učešće nosača zbog olakšane difuzije. Glukoza se tada oslobađa u krvotok. Normalno, pri normalnoj koncentraciji glukoze u krvi i, prema tome, u primarnom urinu, sva glukoza se reapsorbira. Sa viškom glukoze u krvi, što znači da u primarnom urinu može doći do maksimalnog opterećenja tubularnog transportnog sistema, tj. svi molekuli nosači.
U tom slučaju, glukoza se više ne može reapsorbirati i pojavit će se u konačnom urinu (glukozurija). Ovu situaciju karakterizira koncept "maksimalnog cijevastog transporta" (TM). Vrijednost maksimalnog tubularnog transporta odgovara starom konceptu "praga bubrežnog izlučivanja". Za glukozu, ova vrijednost je 10 mmol/l.
Supstance čija reapsorpcija ne ovisi o njihovoj koncentraciji u krvnoj plazmi nazivaju se negranične. To uključuje tvari koje se ili uopće ne reapsorbiraju (inulin, manitol) ili se malo reapsorbiraju i izlučuju urinom srazmjerno njihovom nakupljanju u krvi (sulfati).
Amino kiseline. Reapsorpcija aminokiselina se također odvija mehanizmom Na+-spregnutog transporta. Aminokiseline filtrirane u glomerulima se 90% reapsorbuju u ćelijama proksimalnog tubula bubrega. Ovaj proces se odvija uz pomoć sekundarnog aktivnog transporta, tj. energija ide natrijum pumpi. Postoje najmanje 4 transportna sistema za prijenos različitih aminokiselina (neutralne, dvobazne, dikarboksilne i aminokiseline). Ovi transportni sistemi takođe deluju u crevima za apsorpciju aminokiselina.
tubularna reapsorpcija
Opisani su genetski defekti gdje se određene aminokiseline ne resorbiraju i apsorbiraju u crijevima.
Protein. Normalno, mala količina proteina ulazi u filtrat i ponovo se apsorbuje. Proces reapsorpcije proteina provodi se uz pomoć pinocitoze. Epitel bubrežnih tubula aktivno hvata protein. Po ulasku u ćeliju, protein se hidrolizira enzimima lizosoma i pretvara u aminokiseline. Ne podliježu svi proteini hidrolizi, neki od njih prolaze u krv nepromijenjeni. Ovaj proces je aktivan i zahtijeva energiju. Sa konačnim urinom ne gubi se više od 20-75 mg proteina dnevno. Pojava proteina u urinu naziva se proteinurija. Proteinurija se može pojaviti i u fiziološkim uslovima, na primjer, nakon teškog mišićnog rada. U osnovi, proteinurija se javlja u patologiji nefritisa, nefropatija i multiplog mijeloma.
Urea. Ima važnu ulogu u mehanizmima koncentracije urina, slobodno filtriranog u glomerulima. U proksimalnom tubulu, dio uree se pasivno reapsorbuje gradijentom koncentracije koji nastaje zbog koncentracije urina. Ostatak uree dospijeva u sabirne kanale. U sabirnim kanalićima, pod uticajem ADH, voda se reapsorbuje i povećava se koncentracija uree. ADH povećava propusnost zida za ureu, te ona prelazi u medulu bubrega, stvarajući ovdje otprilike 50% osmotskog tlaka.
Iz intersticija urea difundira duž gradijenta koncentracije u Henleovu petlju i ponovo ulazi u distalne tubule i sabirne kanale. Tako se odvija intrarenalna cirkulacija ureje. U slučaju vodene diureze, apsorpcija vode u distalnom nefronu prestaje, a izlučuje se više uree. Dakle, njegovo izlučivanje zavisi od diureze.
Slabe organske kiseline i baze. Reapsorpcija slabih kiselina i baza zavisi od toga da li su u jonizovanom ili nejonizovanom obliku. Slabe baze i kiseline u jonizovanom stanju se ne reapsorbuju i izlučuju se urinom. Stupanj jonizacije baza se povećava u kiseloj sredini, pa se one brže izlučuju kiselim urinom, slabe kiseline se, naprotiv, brže izlučuju alkalnom urinom.
Ovo je od velike važnosti, jer su mnoge ljekovite tvari slabe baze ili slabe kiseline. Stoga je u slučaju trovanja acetilsalicilnom kiselinom ili fenobarbitalom (slabe kiseline) potrebno primijeniti alkalne otopine (NaHCO3) kako bi se ove kiseline prešle u jonizirano stanje, čime bi se olakšalo njihovo brzo eliminiranje iz organizma. Za brzo izlučivanje slabih baza potrebno je u krv uvesti kisele produkte za zakiseljavanje urina.
Voda i elektroliti. Voda se reapsorbuje u svim dijelovima nefrona. Otprilike 2/3 sve vode se reapsorbira u proksimalnim zavijenim tubulima. Oko 15% se reapsorbira u Henleovoj petlji, a 15% u distalnim izvijenim tubulima i sabirnim kanalima. Voda se pasivno reapsorbuje zbog transporta osmotski aktivnih supstanci: glukoze, aminokiselina, proteina, natrijuma, kalijuma, kalcijuma, jona hlora. Sa smanjenjem reapsorpcije osmotski aktivnih supstanci, smanjuje se i reapsorpcija vode. Prisustvo glukoze u konačnom urinu dovodi do povećanja diureze (poliurije).
Natrijum je glavni ion odgovoran za pasivnu apsorpciju vode. Natrijum je, kao što je već pomenuto, takođe neophodan za transport glukoze i aminokiselina. Osim toga, igra važnu ulogu u stvaranju osmotski aktivnog okruženja u intersticiju bubrežne medule, čime se koncentriše urin. Reapsorpcija natrijuma se javlja u svim dijelovima nefrona. Oko 65% jona natrijuma se reapsorbuje u proksimalnom tubulu, 25% u nefronskoj petlji, 9% u distalnom uvijenom tubulu i 1% u sabirnim kanalima.
Protok natrijuma iz primarnog urina kroz apikalnu membranu u ćeliju tubulnog epitela odvija se pasivno duž elektrohemijskog i koncentracijskog gradijenta. Izlučivanje natrijuma iz ćelije kroz bazolateralne membrane se odvija aktivno uz pomoć Na +, K + - ATPaze. Budući da se energija staničnog metabolizma troši na prijenos natrijuma, njegov transport je primarno aktivan. Transport natrijuma u ćeliju može se odvijati kroz različite mehanizme. Jedna od njih je zamjena Na + za H + (protivstrujni transport, ili antiport). U ovom slučaju, jon natrijuma se prenosi unutar ćelije, a ion vodonika se prenosi van.
Drugi način prijenosa natrijuma u ćeliju provodi se uz sudjelovanje aminokiselina, glukoze. Ovo je takozvani kotransport, ili simport. Djelomično je reapsorpcija natrijuma povezana sa izlučivanjem kalija.
Srčani glikozidi (strofantin K, oubain) su u stanju da inhibiraju enzim Na +, K + - ATPazu, koji osigurava prijenos natrijuma iz stanice u krv i transport kalija iz krvi u ćeliju.
Od velikog značaja u mehanizmima reapsorpcije vode i jona natrijuma, kao i koncentraciji urina, je rad tzv. rotaciono-protivstrujnog multiplikatorskog sistema.
Rotaciono-protivstrujni sistem predstavljen je paralelnim kolenima Henleove petlje i sabirnim kanalom, duž kojeg se tečnost kreće u različitim pravcima (protivstruja). Epitel silaznog dijela petlje je propustljiv za vodu, a epitel uzlaznog koljena je nepropustan za vodu, ali je u stanju da aktivno prenosi ione natrijuma u tkivnu tekućinu, a preko nje natrag u krv. U proksimalnom dijelu, natrijum i voda se apsorbiraju u ekvivalentnim količinama, a urin je ovdje izotoničan u odnosu na krvnu plazmu.
U silaznoj nefronskoj petlji voda se reapsorbuje i urin postaje više koncentriran (hipertoničan). Povrat vode se odvija pasivno zbog činjenice da se u uzlaznom dijelu istovremeno provodi aktivna reapsorpcija natrijevih iona. Ulazeći u tkivnu tečnost, joni natrijuma povećavaju osmotski pritisak u njoj, čime se olakšava privlačenje vode iz silaznog dela u tkivnu tečnost. Istovremeno, povećanje koncentracije urina u nefronskoj petlji zbog reapsorpcije vode olakšava prijelaz natrijuma iz urina u tkivnu tekućinu. Kako se natrijum reapsorbuje u uzlaznom ekstremitetu Henleove petlje, urin postaje hipotoničan.
Ulazeći dalje u sabirne kanale, koji su treće koljeno protustrujnog sistema, urin može biti visoko koncentrisan ako djeluje ADH, što povećava propusnost zidova za vodu. U tom slučaju, kako se krećete duž sabirnih kanala u dubinu medule, sve više vode ulazi u intersticijsku tekućinu, čiji je osmotski tlak povećan zbog sadržaja velike količine Na "1" i uree u to, a urin postaje sve više i više koncentrisan.
Kada velike količine vode uđu u tijelo, bubrezi, naprotiv, luče velike količine hipotonične mokraće.
Tubularna reapsorpcija i izlučivanje tvari u nefronu.
CEVNA REABSORPCIJA ili reverzna apsorpcija u krv vode, soli, organskih supstanci (glukoze, proteina, aminokiselina, vitamina) sadržanih u primarnom urinu.
Rezultat je smanjenje primarnog urina (za 70%), potpuna reapsorpcija u krv tvari korisnih za metabolizam (aminokiseline, glukoza, mnogi vitamini), djelomična apsorpcija vode i jona Na, Cl, K, Ca, izlučivanje toksični metabolički produkti iz krvi u mokraću (urea, mokraćna kiselina, amonijak, kreatinin, sulfati, fosfati).
Apsorpcija baznih supstanci se vrši pomoću mehanizama aktivnog transporta, difuzije i olakšane difuzije.
Na primjer:
Glavni ion koji određuje osmotski pritisak, a samim tim i reapsorpciju vode, Na+ ulazi u epitelne ćelije pasivno, duž gradijenta koncentracije, a zatim se izbacuje sa druge strane ćelije Na + -K + -ATPazom .
K+ joni se aktivno reapsorbuju na apikalnoj membrani i zatim difuzijom puštaju u krv.
Proksimalni izvijeni tubul reapsorbuje 70% vode i jona.
Reapsorpcija kationa (Na+, K+, Ca2+, Mg2+) se odvija protiv gradijenta koncentracije, aktivno (koristeći ATP energiju).
Negativno nabijene anione privlače pozitivno nabijeni kationi, a zbog elektrostatičkih sila pasivno prelaze iz urina u krv (Cl- i HCO3- nakon Na+ i K+; SO42- i PO42- nakon Ca2+ i Mg2+), voda se pasivno apsorbira nakon jona osmotskim gradijentom.
Mehanizmi reapsorpcije Ca2+, Mg2+, SO4-, PO4- slični su mehanizmima reapsorpcije Na+, K+ i Cl-.
Supstance se mogu transportovati u citoplazmu bubrežne epitelne ćelije pomoću nosača zajedno sa Na+ jonima.
Istovremeno, oni ulaze u krv iz epitelne ćelije difuzijom duž gradijenta koncentracije.
Pri određenoj koncentraciji krvnih supstanci (prag izlučivanja), te tvari (prag) neće se potpuno reapsorbirati, a dio filtriranih tvari će završiti u konačnom urinu.
Pražne supstance uključuju glukozu, koja se normalno (4,6-7,2 mmol/l u krvi) filtrira i zatim potpuno reapsorbuje.
S povećanjem njegove koncentracije u krvi na 10,8 mmol / l, dio glukoze neće imati vremena da se reapsorbira.
Izlučuje se urinom iz tijela i dolazi do glukozurije.
REABSORPCIJA u različitim dijelovima nefrona nije ista.
U PROKSIMALNOM ODJELU 40-45% vode, natrijuma, bikarbonata, hlora, aminokiselina, glukoze, vitamina, proteina, mikroelemenata se reapsorbuje do kraja sekcije - 1/3 ultrafiltrata ostaje sa istim osmotskim pritiskom kao u plazmi.
U Henleovoj petlji se reapsorbuje 25-28% vode, do 25% natrijuma, kao i joni hlorida, kalijuma, kalcijuma, magnezijuma
U DISTALNOM ODJELU - 10% vode, oko 9% natrijuma, kalijuma.
U sabirnim tubulima - 20% vode, manje od 1% natrijuma.
TUBULARNA SEKRECIJA se manifestuje ISPUŠTANJEM IZ krvi u LUMINAL CEVICA metaboličkih produkata i stranih supstanci
Tubularna sekrecija je rezultat aktivne aktivnosti epitela bubrežnih tubula.
Izvodi se protiv koncentracijskog ili elektrohemijskog gradijenta i omogućava brzo izlučivanje organskih baza i jona, EPITELNE ĆELIJE luče holin, paraaminohipurnu kiselinu, izmijenjene molekule lijeka iz KRVI iz KRVI i apsorbiraju glutamin iz PRIMARNOG URINA.
Uz pomoć enzima glutaminaze, glutamin se razlaže na GLUTAMINSKA KISELINA i AMONIJAK.
AMONIJAK se izlučuje urinom, koji se izlučuje iz organizma u obliku AMONIJEM SOLI.
Na istom mjestu, ugljičnu kiselinu cijepa enzim UGLJENOHIDRAZA.
Kako teče proces reapsorpcije u bubrezima
HCO3- joni se apsorbuju u krv (zbog elektrostatičke privlačnosti njihovih Na+ i K+).
H+ joni se izlučuju u urinu iz koje se eliminišu.
Ovo objašnjava kiselu reakciju konačnog urina (pH=4,5-6,5).
Ovaj mehanizam Štiti organizam od zakiseljavanja.
LOKALIZACIJA IZLUČIVANJA SUPSTANCI U NEFRONU je drugačija
Ioni vodonika i amonijak se luče u PROKSIMALNOM ODJELU. Osim toga, organske baze se luče u uvijenom dijelu:
Holin, serotonin, dopamin, kinin, morfijum.
U direktnom dijelu - organske kiseline: paraaminohipurna, Diodrast, Penicilin, Mokraćna kiselina.
U DISTALNOM ODJELU - para-aminohipurna kiselina, amonijak, H+ i K+ joni.
LJEKOVITE SUPSTANCE se izlučuju iz organizma uz pomoć GLOMER FILTRACIJE (levomicetin, streptomicin, tetraciklin, neomicin, kanamicin i drugi antibiotici).
Uz pomoć tubularne sekrecije penicilin se izlučuje (za 80-90%).
kada su različiti delovi nefrona oštećeni, određeni broj lekovitih jedinjenja cirkuliše u krvi dugo vremena i ne može se izlučiti iz organizma.
U ovim slučajevima, NEOPHODNO je promijeniti doze LJEKOVIH supstanci.