 Вход
ВходКанальцевая реабсорбция и транспорт основных минеральных веществ. Механизмы реабсорбции в канальцах
text_fields
text_fields
arrow_upward
Сравнение состава и количества первичной и конечной мочи показывает, что в канальцах нефрона происходит процесс обратного всасывания воды и веществ, профильтровавшихся в клубочках. Этот процесс называется каналъцевой реабсорбцией
В зависимости от отдела канальцев, где он происходит, различают реабсорбцию проксимальную и дистальную .
Реабсорбция представляет собой транспорт веществ из мочи в лимфу и кровь и в зависимости от механизма транспорта выделяют пассивную, первично и вторично активную реабсорбцию.
Проксимальная реабсорбция
text_fields
text_fields
arrow_upward
Проксимальная реабсорбция обеспечивает полное всасывание ряда веществ первичной мочи - глюкозы, белка, аминокислот и витаминов. В проксимальных отделах всасывается 2/3 профильтровавшихся воды и натрия, большие количества калия, двухвалентных катионов, хлора, бикарбоната, фосфата, а также мочевая кислота и мочевина. К концу проксимального отдела в его просвете остается только 1/3 объема ультрафильтрата, и, хотя его состав уже существенно отличается от плазмы крови, осмотическое давление первичной мочи остается таким же, как в плазме.
Всасывание воды происходит пассивно, по градиенту осмотического давления и зависит от реабсорбции натрия и хлорида. Реабсорбция натрия в проксимальном отделе осуществляется как активным, так и пассивным транспортом. В начальном участке канальцев это активный процесс. Хотя натрий входит в клетки эпителия через апикальную мембрану пассивно через натриевые каналы по концентрационному и электрохимическому градиенту, его выведение через базолатеральные мембраны эпителиальных клеток происходит активно с помощью натрий-калиевых насосов, использующих энергию АТФ. Сопровождающим всасывающийся натрий анионом является здесь бикарбонат, а хлориды всасываются плохо. Объем мочи в канальце уменьшается из- за пассивной реабсорбции воды, и концентрация хлоридов в его содержимом растет. В конечных участках проксимальных канальцев межклеточные контакты высоко проницаемы для хлоридов (концентрация которых повысилась) и они пассивно по градиенту всасываются из мочи. Вместе с ними пассивно реабсорбируются натрий и вода. Такой пассивный транспорт одного иона (натрия) вместе с пассивным транспортом другого (хлорида) носит название котранспорта.
Таким образом, в проксимальном отделе нефрона существуют два механизма всасывания воды и ионов:
1) активный транспорт натрия с пассивной реабсорбцией бикарбоната и воды,
2) пассивный транспорт хлоридов с пассивной реабсорбцией натрия и воды.
Поскольку натрий и другие электролиты всегда всасываются в проксимальных канальцах с осмотически эквивалентным количеством воды, моча в проксимальных отделах нефрона остается изоосмотичной плазме крови.
Проксимальная реабсорбция глюкозы и аминокислот осуществляется с помощью специальных переносчиков щеточной каемки апикальной мембраны эпителиальных клеток. Эти переносчики транспортируют глюкозу или аминокислоту только если одновременно связывают и переносят натрий. Пассивное перемещение натрия по градиенту внутрь клеток ведет к прохождению через мембрану и переносчика с глюкозой или аминокислотой. Для реализации этого процесса необходима низкая концентрация в клетке натрия, создающая градиент концентрации между внешней и внутриклеточной средой, что обеспечивается энергозависимой работой натрий-калиевого насоса базальной мембраны. Поскольку перенос глюкозы или аминокислоты связан с натрием, а его транспорт определяется активным удалением натрия из клетки, такой вид транспорта называют вторично активным или симпортом, т.е. совместным пассивным транспортом одного вещества (глюкоза) из-за активного транспорта другого (натрия) с помощью одного переносчика.
Поскольку для реабсорбции глюкозы необходимо связывание каждой ее молекулы с молекулой переносчика, очевидно, что при избытке глюкозы может произойти полная загрузка всех молекул переносчиков и глюкоза уже не сможет всасываться в кровь. Эта ситуация характеризуется понятием «максимальный канальцевый транс порт вещества», которое отражает максимальную загрузку канальцевых переносчиков при определенной концентрации вещества в первичной моче и, соответственно, в крови. Постепенно повышая содержание глюкозы в крови и тем самым в первичной моче, можно легко обнаружить ту величину ее концентрации, при которой глюкоза появляется в конечной моче и когда ее экскреция начинает линейно зависеть от прироста уровня в крови. Эта концентрация глюкозы в крови и, соответственно, ультрафильтрате свидетельствует о том, что все канальцевые переносчики достигли предела функциональных возможностей и полностью загружены. В это время реабсорбция глюкозы максимальна и составляет от 303 мг/мин у женщин и до 375 мг/мин у мужчин. Величине максимального канальцевого транспорта соответствует более старое понятие «почечный порог выведения».
Почечным порогом выведения называют ту концентрацию вещества в крови и в первичной моче, при которой оно уже не может быть полностью реабсорбировано в канальцах и появляется в конечной моче.
Такие вещества, для которых может быть найден порог выведения, т.е. реабсорбирующиеся при низких концентрациях в крови полностью, а при повышенных концентрациях - не полностью, носят название пороговых. Типичным примером является глюкоза, которая полностью всасывается из первичной мочи при концентрациях в плазме крови ниже 10 моль/л, но появляется в конечной моче, т.е. полностью не реабсорбируется, при содержании ее в плазме крови выше 10 моль/л. Следовательно, для глюкозы порог выведения составляет 10 моль/л.
Вещества, которые вообще не реабсорбируются в канальцах (инулин, маннитол) или мало реабсорбируются и выделяются пропорционально накоплению в крови (мочевина, сульфаты и др.), называются непороговыми, т.к. для них порога выведения не существует.
Малые количества профильтровавшегося белка практически полностью реабсорбируются в проксимальных канальцах с помощью пиноцитоза. Мелкие белковые молекулы абсорбируются на поверхности апикальной мембраны эпителиальных клеток и поглощаются ими с образованием вакуолей, которые передвигаясь сливаются с лизосомами. Протеолитические ферменты лизосом расщепляют поглощенный белок, после чего низкомолекулярные фрагменты и аминокислоты переносятся в кровь через базолатеральную мембрану клеток.
Дистальная реабсорбция
text_fields
text_fields
arrow_upward
Дистальная реабсорбция ионов и воды по объему значительно меньше проксимальной. Однако, существенно меняясь под влиянием регулирующих воздействий, она определяет состав конечной мочи и способность почки выделять либо концентрированную, либо разведенную мочу (в зависимости от водного баланса организма). В дистальном отделе нефрона происходит активная реабсорбция на трия. Хотя здесь всасывается всего 10% от профильтровавшегося количества катиона, этот процесс обеспечивает выраженное уменьшение его концентрации в моче и, напротив, повышение концентрации в интерстициальной жидкости, что создает значительный градиент осмотического давления между мочой и интерстицием. Хлор всасывается преимущественно пассивно вслед за натрием. Способность эпителия дистальных канальцев секретировать в мочу Н-ионы связана с реабсорбцией ионов натрия, этот вид транспорта в виде обмена натрия на протон получил название «антипорт». Активно всасывается в дистальном отделе канальцев калий, кальций и фос фаты. В собирательных трубочках, главным образом юкстамедуллярных нефронов, под влиянием вазопрессина повышается проницаемость стенки для мочевины и она, благодаря высокой концентрации в просвете канальца, пассивно диффундирует в окружающее интерстициальное пространство, увеличивая его осмолярность. Под влиянием вазопрессина стенка дистальных извитых канальцев и собирательных трубочек становится проницаемой и для воды, в результате чего происходит ее реабсорбция по осмотическому градиенту в гиперосмолярный интерстиций мозгового вещества и далее в кровь.
Способность почки образовывать концентрированную или разведенную мочу обеспечивается деятельностью противоточно-множи тельной канальцевой системы почки, которая представлена параллельно расположенными коленами петли Генле и собирательными трубочками (рис.12.2).
Цифрами обозначены величины осмотического давления интерстициальной жидкости и мочи. В собирательной трубочке цифрами в скобках обозначено осмотическое давление мочи в отсутствие вазопрессина (разведение мочи), цифрами без скобок - осмотическое давление мочи в условиях действия вазопрессина (концентрирование мочи).
Моча двигается в этих канальцах в противоположных направлениях (почему систему и назвали противоточной), а процессы транспорта веществ в одном колене системы усиливаются («умножаются») за счет деятельности другого колена. Определяющую роль в работе противоточного механизма играет восходящее колено петли Генле, стенка которого непроницаема для воды, но активно реабсорбирует в окружающее интерстициальное пространство ионы натрия. В результате, интерстициальная жидкость становится гиперосмотичной по отношению к содержимому нисходящего колена петли и по направлению к вершине петли осмотическое давление в окружающей ткани растет. Стенка же нисходящего колена проницаема для воды, которая пассивно уходит из просвета в гиперосмотичный интерстиций. Таким образом, в нисходящем колене моча из-за всасывания воды становится все более и более гиперосмотичной, т.е. устанавливается осмотическое равновесие с интерстициальной жидкостью. В восходящем колене, из-за всасывания натрия, моча становится все менее осмотичной и в корковый отдел дистального канальца восходит уже гипотоничная моча. Однако ее количество из-за всасывания воды и солей в петле Генле существенно уменьшилось.
Собирательная трубочка, в которую затем поступает моча, тоже образует с восходящим коленом петли Генле противоточную систему. Стенка собирательной трубочки становится проницаемой для воды только в присутствии вазопрессина. В этом случае, по мере продвижения мочи по собирательным трубочкам вглубь мозгового вещества, в котором нарастает осмотическое давление из-за всасывания натрия в восходящем колене петли Генле, все больше воды пассивно уходит в гиперосмотичный интерстиций и моча становится все более концентрированной.
Под влиянием вазопрессина реализуется еще один важный для концентрирования мочи механизм - пассивный выход мочевины из собирательных трубочек в окружающий интерстиций. Всасывание воды в верхних отделах собирательных трубочек ведет к нарастанию концентрации мочевины в моче, а в самых нижних их отделах, расположенных в глубине мозгового вещества, вазопрессин повышает проницаемость для мочевины и она пассивно диффундирует в интерстиций, резко повышая его осмотическое давление. Таким образом, интерстиций мозгового вещества становится наиболее высоко осмотичным в области вершины почечных пирамид, где и происходит увеличение всасывания воды из просвета канальцев в интерстиций и концентрирование мочи.
Мочевина интерстициальной жидкости по концентрационному градиенту диффундирует в просвет тонкой восходящей части петли Генле и вновь поступает с током мочи в дистальные канальцы и собирательные трубочки. Так осуществляется кругооборот мочевины в канальцах, сохраняющих высокий уровень ее концентрации в мозговом веществе. Описанные процессы протекают в основном в юкстамедуллярных нефронах, имеющих наиболее длинные петли Генле, спускающиеся глубоко внутрь мозгового вещества почки.
В мозговом веществе почки имеется и другая - сосудистая про тивоточная система, образованная кровеносными капиллярами. Поскольку кровеносная сеть юкстамедуллярных нефронов образует длинные параллельные прямые нисходящие и восходящие капиллярные сосуды (рис. 12.1), спускающиеся вглубь мозгового вещества, двигающаяся по нисходящему прямому капиллярному сосуду кровь постепенно отдает воду в окружающее интерстициальное пространство в силу нарастающего осмотического давления в ткани и, напротив, обогащается натрием и мочевиной, сгущается и замедляет свое движение. В восходящем капиллярном сосуде по мере движения крови в ткани с постепенно снижающимся осмотическим давлением происходят обратные процессы - натрий и мочевина по концентрационному градиенту диффундируют обратно в ткань, а вода всасывается в кровь. Таким образом, и эта противоточная система способствует поддержанию высокого осмотического давления в глубоких слоях ткани мозгового вещества, обеспечивая удаление воды и удержание натрия и мочевины в интерстиций.
Деятельность описанных противоточных систем во многом зависит от скорости движения находящихся в них жидкостей (мочи или крови). Чем скорее будет двигаться моча по трубкам противоточной системы канальцев, тем меньшие количества натрия, мочевины и воды успеют реабсорбироваться в интерстиций и большие количества менее концентрированной мочи будут выделяться почкой. Чем выше будет скорость кровотока по прямым капиллярным сосудам мозгового вещества почки, тем больше натрия и мочевины унесет кровь из почечного интерстиция, т.к. они не успеют диффундировать из крови назад в ткань. Этот эффект называют «вымыванием» осмотически активных веществ из интерстиция, в результате его осмолярность падает, концентрирование мочи уменьшается и почкой выделяется больше мочи низкого удельного веса (разведение мочи). Чем медленнее происходит движение мочи или крови в мозговом веществе почек, тем больше осмотически активных веществ накапливается в интерстиции и выше способность почки концентрировать мочу.
Регуляция каналъцевой реабсорбции
text_fields
text_fields
arrow_upward
Регуляция каналъцевой реабсорбции осуществляется как нервным , так и, в большей мере, гуморальным путем.
Нервные влияния преимущественно реализуются симпатическими проводниками и медиаторами через бета- адренорецепторы мембран клеток проксимальных и дистальных канальцев. Симпатические эффекты проявляются в виде активации процессов реабсорбции глюкозы, натрия, воды и фосфатов и реализуются через систему вторичных посредников (аденилатциклаза - цАМФ). В регуляции процессов метаболизма почечной ткани существенную роль играют трофические влияния симпатической нервной системы. Нервная регуляция кровообращения в мозговом веществе почки увеличивает или уменьшает эффективность сосудистой противоточной системы и концентрирование мочи.
Сосудистые эффекты нервной регуляции могут опосредоваться через внутрипочечные системы гуморальных регуляторов - ренин- ангиотензинную, кининовую, простагландины и др. Основным фактором регуляции реабсорбции воды в дистальных отделах нефрона является гормон вазопрессин, называвшийся ранее антидиуретическим гормоном. Этот гормон образуется в супраоптическом и паравентрикулярных ядрах гипоталамуса и поступает в кровь из нейрогипофиза. Влияние вазопрессина на проницаемость эпителия канальцев обусловлено наличием рецепторов к гормону, относящихся к V-2 типу, на поверхности базолатеральной мембраны клеток эпителия. Образование гормон-рецепторного комплекса (глава 3), влечет за собой через посредство GS-белка и гуанилового нуклеотида активацию аденилатциклазы и образование цАМФ у базолатеральной мембраны (рис. 12.3).
Рис. 12.3. Механизм действия вазопрессина на проницаемость собирательных трубочек для воды.Рис. 12.3. Механизм действия вазопрессина на проницаемость собирательных трубочек для воды.
Б-л мембрана - базолатеральная мембрана клеток,
А мембрана - апикальная мембрана,
ГН - гуанидиновый нуклеотид,АЦ - аденилатциклаза.
После этого цАМФ пересекает клетку эпителия и, достигнув апикальной мембраны, активирует цАМФ- зависимые протеинкиназы. Под влиянием этих ферментов происходит фосфорилирование мембранных белков, приводящее к повышению проницаемости для воды и увеличению поверхности мембраны. Перестройка ультраструктур клетки ведет к образованию специализированных вакуолей, переносящих большие потоки воды по осмотическому градиенту от апикальной к базолатеральной мембране, не позволяя самой клетке набухать. Такой транспорт воды через клетки эпителия реализуется вазопрессином в собирательных трубочках. Кроме того, в дистальных канальцах вазопрессин обусловливает активацию и выход из клеток гиалуронидаз, вызывающих расщепление гликозаминогликанов основного межклеточного вещества и межклеточный пассивный транспорт воды по осмотическому градиенту.
Канальцевая реабсорбция воды
text_fields
text_fields
arrow_upward
Канальцевая реабсорбция воды регулируется и другими гормонами.
С учетом механизмов действия все гормоны, регулирующие реабсорбцию воды, можно представить в виде шести групп:
1) повышающие проницаемость мембран дистальных отделов нефрона для воды (вазопрессин, пролактин, хорионический гонадотропин);
2) меняющие чувствительность клеточных рецепторов к вазопрессину (паратирин, кальцитонин, кальцитриол, простагландины, альдостерон);
3) меняющие осмотический градиент интерстиция мозгового слоя почки и, соответственно, пассивный осмотический транспорт воды (паратирин, кальцитриол, тиреоидные гормоны, инсулин, вазопрессин);
4) меняющие активный транспорт натрия и хлорида, а за счет этого и пассивный транспорт воды (альдостерон, вазопрессин, атриопептид, прогестерон, глюкагон, кальцитонин, простагландины);
5) повышающие осмотическое давление канальцевой мочи за счет нереабсорбированных осмотически активных веществ, например глюкозы (контринсулярные гормоны);
6) меняющие кровоток по прямым сосудам мозгового.вещества и, тем самым, накопление или «вымывание» осмотически активных веществ из интерстиция (ангиотензин- II, кинины, простагландины, паратирин, вазопрессин, атриопептид).
Канальцевая реабсорбция электролитов
text_fields
text_fields
arrow_upward
Канальцевая реабсорбция электролитов, также как и воды, регулируется преимущественно гормональными, а не нервными влияниями.
Реабсорбция натрия в проксимальных канальцах активируется альдостероном и угнетается паратирином, в толстой части восходящего калена петли Генле реабсорбция натрия активируется вазопрессином, глюкагоном, кальцитонином, а угнетается простагландинами Е. В дистальном отделе канальцев основными регуляторами транспорта натрия являются альдостерон (активация), простагландины и атриопептид (угнетение).
Регуляция канальцевого транспорта кальция, фосфата и частично магния обеспечивается, в основном, кальций-регулирующими гормонами. Паратирин имеет в канальцевом аппарате почки несколько участков действия. В проксимальных канальцах (прямой отдел) всасывание кальция происходит параллельно с транспортом натрия и воды. Угнетение реабсорбции натрия в этом отделе под влиянием паратирина сопровождается параллельным снижением реабсорбции кальция. За пределами проксимального канальца паратирин избирательно усиливает реабсорбцию кальция, особенно в дистальном извитом канальце и корковой части собирательных трубочек. Реабсорбция кальция активируется также кальцитриолом, а подавляется кальцитонином. Всасывание фосфата в канальцах почки угнетается и паратирином (проксимальная реабсорбция), и кальцитонином (дистальная реабсорбция), а усиливается кальцитриолом и соматотропином. Паратирин активирует реабсорбцию магния в корковой части восходящего колена петли Генле и тормозит проксимальную реабсорбцию бикарбоната.
В организм человека и выведение продуктов обмена осуществляет выделительная система человека. Работа органов выделительной системы человека имеет свои сформированные в процессе эволюции механизмы выведения продуктов обмена, которыми являются фильтрация, реабсорбция и секреция.
Выделительная система человека
Выведение продуктов обмена из организма осуществляют которые состоят из почек, мочеточников, мочевого пузыря и мочеиспускательного канала.
Почки расположены в забрюшинном пространстве в области поясничного отдела и имеют бобовидную форму.
Это парный орган, состоящий из коркового и мозгового вещества, лоханки, и покрыта она фиброзной оболочкой. Лоханка почки состоит из малой и большой чаши, и из нее выходит мочеточник, который доставляет мочу в мочевой пузырь и через мочеиспускательный канал конечная моча выводится из организма.
Почки участвуют в обменных процессах, и их роль в обеспечении водного баланса организма, поддержании кислотно-щелочного баланса являются основополагающей для полноценного существования человека.
Строение почки очень сложное и ее структурным элементом является нефрон.

Он имеет сложное строение и состоит из проксимального канала, тельца нефрона, петли Генле, дистального канала и собирательной трубочки, дающей начало мочеточникам. Реабсорбция в почках проходит через канальцы проксимальной, дистальной части и петли Генле.
Механизм реабсорбции
Молекулярные механизмы прохождения веществ в процессе реабсорбции это:
- диффузия;
- эндоцитоз;
- пиноцитоз;
- пассивный транспорт;
- активный транспорт.
Особое значение имеет для реабсорбции активный и пассивный транспорт и направление реабсорбируемых веществ по электрохимическому градиенту и наличию переносчика для веществ, работа клеточных насосов и другие характеристики.
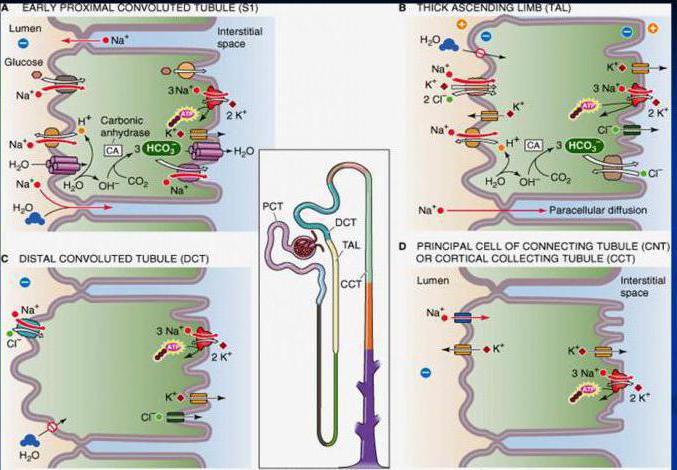
Веществ идет против электрохимического градиента с затратой энергии на ее реализацию и через специальные транспортные системы. Характер передвижения - трансцеллюлярный, который осуществляется переходом через апикальную мембрану и базолатеральную. Такими системами являются:
- Первично-активный транспорт, который осуществляется с помощью энергии от расщепления АТФ. Его используют ионы Na+, Ca+, K+, H+.
- Вторично-активный транспорт, проходит за счет разницы в концентрации ионов натрия в цитоплазме и в просвете канальцев, и эта разница объясняется выходом ионов натрия в межклеточную жидкость с затратой энергии расщепления АТФ. Его используют аминокислоты, глюкоза.
Проходит по градиентам: электрохимическому, осмотическому, концентрационному, и для его осуществления не требуется затрат энергии и образования переносчика. Вещества, которые используют его - это ионы Cl-. Движение веществ осуществляется парацеллюлярно. Это движение через мембрану клетки, которая расположена между двумя клетками. Характерными молекулярными механизмами являются диффузия, перенос с растворителем.
Процесс реабсорбции белка проходит внутри клеточной жидкости, и, после расщепления его на аминокислоты, они поступают в межклеточную жидкость, что происходит в результате пиноцитоза.
Виды реабсорбции
Реабсорбция - это процесс, проходящий в канальцах. И вещества, проходящие через канальцы, имеют разные переносчики и механизмы.

В сутки в почках формируется от 150 до 170 литров первичной мочи, которая проходит процесс реабсорбции и возвращается в организм. Вещества, имеющие высокодисперсные компоненты, не могут пройти через мембрану канальцев и в процессе реабсорбции поступают в кровь с другими веществами.
Проксимальная реабсорбция
В проксимальном отделе нефрона, который расположен в корковом веществе почки, реабсорбция проходит для глюкозы, натрия, воды, аминокислот, витаминов и белка.

Проксимальный каналец образован эпителиальными клетками, которые имеют апикальную мембрану и щеточную каемку, и обращена она в сторону просвета почечных канальцев. Базальная мембрана образует складки, формирующие базальный лабиринт, и через них первичная моча попадает в перитубулярные капилляры. Клетки между собой соединены плотно и образуют пространство, которое проходит на всем протяжении межклеточного пространства канальца, и называется он базолатеральным лабиринтом.
Реабсорбция натрия имеет сложный трехступенчатый этап, и он является переносчиком для других веществ.
Реабсорбция ионов, глюкозы и аминокислот в проксимальном канальце
Основные этапы реабсорбции натрия:
- Прохождение через апикальную мембрану. Это - этап пассивного транспорта натрия, через Na-каналы и Na-переносчиков. Ионы натрия проходят в клетку через мембранные гидрофильные белки, образующие Na-каналы.
- Поступление или прохождение через мембрану сопряжено с обменом Na+ на водород, например, или же с поступлением его как переносчика глюкозы, аминокислоты.
- Прохождение через базальную мембрану. Это - этап активного транспорта Na+, через Na+/K+ насосы с помощью фермента АТФ, который при расщеплении выделяет энергию. Натрий, реабсорбируясь в почечных канальцах, постоянно возвращается в обменные процессы и его концентрация в клетках проксимального канальца - низкая.
Реабсорбция глюкозы проходит по вторично-активному транспорту и ее поступление облегчено за счет переноса ее через Na-насос, и она полностью возвращается в обменные процессы в организме. Повышенная концентрация глюкозы не проходит полностью реабсорбцию в почках и выделяется с конечной мочой.

Реабсорбция аминокислот проходит аналогично глюкозе, но сложная организация аминокислот требует участия специальных переносчиков для каждой аминокислоты на менее 5-7 дополнительных.
Реабсорбция в петле Генле
Петля Генле проходит через и процесс реабсорбции в восходящей и нисходящей части ее для воды и ионов отличается.
Фильтрат, попадая в нисходящую часть петли, спускаясь по ней, отдает воду за счет разного градиента давления и насыщается ионами натрия и хлора. В этой части вода реабсорбируется, а для ионов она непроницаема. Восходящая часть непроницаема для воды и при прохождении через нее первичная моча разбавляется, тогда, как в нисходящей концентрируется.
Дистальная реабсорбция
Этот отдел нефрона расположен в корковом веществе почки. Его функция заключается в реабсорбции воды, которая собирается с первичной мочой и подвергает реабсорбции ионы натрия. Дистальная реабсорбция - это разведение первичной мочи и формирование из фильтрата мочи конечной.
Поступая в дистальный каналец, первичная моча в объеме 15% после реабсорбции в почечных канальцах, составляет 1% общего объема. Собираясь после этого в собирательной трубочке, она разбавляется, и формируется конечная моча.
Нейро-гуморальная регуляция реабсорбции
Реабсорбция в почках регулируется симпатической нервной системой и гормонами щитовидной железы, гипоталамо-гипофизарными и андрогенами.
Реабсорбция натрия, воды, глюкозы увеличивается при возбуждении симпатических и блуждающих нервов.
Дистальные канальца и собирательные трубочки осуществляют реабсорбцию воды в почках под влиянием антидиуретического гормона или вазопрессина, который при уменьшении воды в организме увеличивается в больших количествах, а также увеличивается проницаемость стенок канальцев.
Альдостерон увеличивает реабсорбцию кальция, хлора и воды, так же, как и атриопептид, который вырабатывается в правом предсердии. Угнетение реабсорбции натрия в проксимальном отделе нефрона идет при поступлении паратирина.
Активация реабсорбции натрия идет за счет гормонов:
- Вазопрессин.
- Глюкоган.
- Кальцитонином.
- Альдостероном.
Угнетение реабсорбции натрия идет при выработке гормонов:
- Простагландин и простагландин Е.
- Атриопептид.
Кора головного мозга осуществляет регуляцию выведения или затормаживания мочи.
Канальцевая реабсорбция воды осуществляется множеством гормонов, отвечающих за проницаемость мембран дистального отдела нефрона, регуляцию транспорта ее по канальцам и многое другое.
Значение реабсорбции
Практическое применение научных знаний о том, что такое реабсорбция - это в медицине позволило получить информационное подтверждение о работе выделительной системы организма и заглянуть во внутренние его механизмы. проходит очень сложные механизмы и влияние на него окружающей среды, генетических отклонений. И они не остаются незамеченными при возникновении проблем на их фоне. Одним словом, здоровье - это очень важно. Следите за ним и за всеми процессами, происходящими в организме.
Подробности
Реабсорбция – это транспорт веществ из просвета почечных канальцев в кровь , протекающую через околоканальцевые капилляры. Реабсорбируется 65% от объема первичной мочи (примерно 120 л/сутки. Было 170 л, выделилось 1.5): вода, минеральные соли, все необходимые органические компоненты, (глюкоза, аминокислоты). Транспорт пассивный (осмос, диффузия по электрохимическому градиенту) и активный (первично-активный и вторично-активный с участием белковых молекул-переносчиков). Транспортные системы такие же, как и в тонком кишечнике.
Пороговые вещества – обычно полностью реабсорбируются
(глюкоза, аминокислоты) и выделяются с мочой только если их концентрация в плазме крови превышает пороговую величину (так называемый «порог выведения»). Для глюкозы порог выведения 10 ммоль/л (при нормальной концентрации глюкозы в крови 4.4-6.6 ммоль/л).
Беспороговые вещества – всегда выводятся независимо от их концентрации в плазме крови . Они не реабсорбируются или реабсорбируются частично, например, мочевина и др. метаболиты.
Механизм работы различных отделов почечного фильтра.
1. В проксимальном канальце берет свое начало процесс концентрирования клубочкового фильтрата, причем наиболее важным моментом здесь является активное поглощение солей. С помощью активного транспорта из данного участка канальца обратно всасывается около 67% Na+. Почти пропорциональное количество воды и некоторых других растворенных веществ, например ионов хлора, следует за ионами натрия пассивно. Таким образом, прежде чем фильтрат достигнет петли Генле, из него реабсорбирустся около 75% веществ. В результате канальцевая жидкость становится изоосмотической по отношению к плазме крови и тканевым жидкостям.
Проксимальный каналец идеально приспособлен для интенсивной реабсорбции соли и воды . Многочисленные микроворсинки эпителия образуют так называемую щеточную кайму, покрывающую внутреннюю поверхность просвета почечного канальца. При таком устройстве абсорбирующей поверхности чрезвычайно увеличивается площадь клеточной мембраны и в результате облегчается диффузия соли и воды из просвета канальца в эпителиальные клетки.
2. Нисходящее колено петли Генле и часть восходящего колена , расположенная во внутреннем слое мозгового вещества , состоят из очень тонких клеток, у которых нет щеточной каймы, а число митохондрий мало. Морфология тонких участков нефрона свидетельствует об отсутствии здесь активного переноса растворенных веществ через стенку канальца. На данном участке нефрона NaCl очень плохо проникает сквозь стенку канальца, мочевина - несколько лучше, а вода проходит без затруднений.
3. Стенка тонкого участка восходящего колена петли Генле также неактивна в отношении транспорта соли. Тем не менее она обладает высокой проницаемостью для Na+ и Сl-, но малопроницаема для мочевины и почти непроницаема для воды.
4. Толстый участок восходящего колена петли Генле , расположенный в мозговом веществе почки, отличается от остальных участков указанной петли. Он осуществляет активный перенос Na+ и Cl- из просвета петли в интерстициальное пространство. Этот участок нефрона вместе с остальной частью восходящего колена чрезвычайно мало проницаем для воды. Из-за реабсорбции NaCl жидкость поступает в дистальный каналец несколько гипоосмотичной по сравнению с тканевой жидкостью
5. Движение воды через стенку дистального канальца - процесс сложный. Дистальный каналец имеет особое значение для транспорта К+, Н+ и NH3 из тканевой жидкости в просвет нефрона и транспорта Na+, Cl- и Н2О из просвета нефрона в тканевую жидкость. Поскольку соли активно "выкачиваются" из просвета канальца, вода следует за ними пассивно.
6. Собирательный проток проницаем для воды, что позволяет ей переходить из разбавленной мочи в более концентрированную тканевую жидкость мозгового вещества почки. В этом заключается конечная стадия образования гиперосмотической мочи. В протоке происходит также реабсорбция NaCl, но за счет активного переноса Na+ через стенку. Для солей собирательный проток непроницаем, в отношении воды его проницаемость меняется. Важной особенностью дистального участка собирательного протока, расположенного во внутреннем мозговом слое почек, является его высокая проницаемость для мочевины.

Механизм реабсорбции глюкозы.
Проксимальная (1/3) реабсорбция глюкозы осуществляется с помощью специальных переносчиков щеточной каемки апикальной мембраны эпителиальных клеток . Эти переносчики транспортируют глюкозу, только если одновременно связывают и переносят натрий. Пассивное перемещение натрия по градиенту концентрации внутрь клеток ведет к транспорту через мембрану и переносчика с глюкозой.
Для реализации этого процесса необходима низкая концентрация натрия в эпителиальной клетке, создающая градиент концентрации между внешней и внутриклеточной средой, что обеспечивается энергозависимой работой натрий-калиевого насоса базальной мембраны .
Такой вид транспорта называют вторично активным, или симпортом , т. е. совместным пассивным транспортом одного вещества (глюкоза) из-за активного транспорта другого (натрия) с помощью одного переносчика. При избытке глюкозы в первичной моче может произойти полная загрузка всех молекул переносчиков и глюкоза уже не сможет всасываться в кровь.
Эта ситуация характеризуется понятием «максимальный канальцевый транспорт вещества » (Тм глюкозы), которое отражает максимальную загрузку канальцевых переносчиков при определенной концентрации вещества в первичной моче и, соответственно, в крови. Эта величина составляет от 303 мг/мин у женщин до 375 мг/мин у мужчин. Величине максимального канальцевого транспорта соответствует понятие «почечный порог выведения».
Почечным порогом выведения называют ту концентрацию вещества в крови и, соответственно, в первичной моче, при которой оно уже не может быть полностью реабсорбировано в канальцах и появляется в конечной моче. Такие вещества, для которых может быть найден порог выведения, т. е. реабсорбирующиеся при низких концентрациях в крови полностью, а при повышенных концентрациях - не полностью, носят название пороговых. Примером является глюкоза, которая полностью всасывается из первичной мочи при концентрациях в плазме крови ниже 10 ммоль/л, но появляется в конечной моче, т. е. полностью не реабсорбируется, при содержании ее в плазме крови выше 10 ммоль/л. Следовательно, для глюкозы порог выведения составляет 10 ммоль/л .

Механизмы секреции в почечном фильтре.
Секреция - это транспорт веществ из крови , протекающей через околоканальцевые капилляры, в просвет почечных канальцев. Транспорт пассивный и активный. Секретируются ионы Н+, К+, аммиак, органические кислоты и основания (например, чужеродные вещества, в частности, лекарственные препараты: пенициллин и др). Секреция органических кислот и оснований происходит с помощью вторично-активного натрий-зависимого механизма.

Секреция йонов калия.
Большая часть легко фильтрующихся в клубочках ионов калия обычно реабсорбируется из фильтрата в проксимальных канальцах и петлях Генлe . Скорость активной реабсорбции в канальце и петле не снижается даже в том случае, когда концентрация К+ в крови и фильтрате сильно возрастает в ответ на избыточное потребление организмом этого иона.
Однако дистальные канальцы и собирательные протоки способны не только реабсорбировать, но и секретировать ионы калия . Секретируя калий, данные структуры стремятся достичь ионного гомеостаза в случае поступления в организм необычайно большого количества этого металла. Транспорт К+, по-видимому, зависит от его постуления в клетки канальцев из тканевой жидкости, обусловленного активностью обычного Nar+ - Ka+-насоса, с утечкой К+ из цитоплазмы в канальцевую жидкость. Калий может просто диффундировать по электрохимическому градиенту из клеток почечных канальцев в просвет, потому что канальцевая жидкость электроотрицательна по отношению к цитоплазме. Секреция К+ с помощью данных механизмов стимулируется адренокортикальным гормоном-альдостероном, который высвобождается в ответ на повышение содержания К+ в плазме крови.
Канальцевая реабсорбция - это процесс обратного всасывания воды и веществ из содержащейся в просвете канальцев мочи в лимфу и кровь.
Основная масса молекул реабсорбируется в проксимальном отделе нефрона. Здесь практически полностью абсорбируются аминокислоты, глюкоза, витамины, белки, микроэлементы, значительное количество ионов Na+, C1-, HCO3- и многие другие вещества.
В петле Генле, дистальном отделе канальца и собирательных трубочках всасываются электролиты и вода.
Альдостерон стимулирует реабсорбцию Na+ и экскрецию K+ и H+ в почечные канальцы в дистальном отделе нефрона, в дистальном канальце и кортикальных собирательных трубочках .
Вазопрессин способствует реабсорбции воды из дистальных извитых канальцев и собирательных трубок.
С помощью пассивного транспорта осуществляется реабсорбция воды, хлора, мочевины.
Активным транспортом называют перенос веществ против электрохимического и концентрационного градиентов. Причем различают первично-активный и вторично-активный транспорт. Первично-активный транспорт происходит с затратой энергии клетки. Примером служит перенос ионов Na+ с помощью фермента Na+/K+-АТФазы, использующей энергию АТФ. При вторично-активном транспорте перенос вещества осуществляется за счет энергии транспорта другого вещества. Механизмом вторично-активного транспорта реабсорбируются глюкоза и аминокислоты.
Величине максимального канальцевого транспорта соответствует старое понятие "почечный порог выведения". Для глюкозы эта величина составляет 10 ммоль/л.
Вещества, реабсорбция которых не зависит от их концентрации в плазме крови, называются непороговыми. К ним относятся вещества, которые или вообще не реабсорбируются, (инулин, маннитол) или мало реабсорбируются и выделяются с мочой пропорционально накоплению их в крови (сульфаты).
В норме небольшое количество белка попадает в фильтрат и реабсорбируется. Процесс реабсорбции белка осуществляется с помощью пиноцитоза. Войдя в клетку, белок подвергается гидролизу со стороны ферментов лизосом и превращается в аминокислоты. Не все белки подвергаются гидролизу, часть их переходит в кровь в неизмененном виде. Этот процесс активный и требует энергии. Появление белка в моче носит название протеинурии. Протеинурия может быть и в физиологических условиях, пример, после тяжелой мышечной работы. В основном протеинурия имеет место в патологии при нефритах, нефропатиях, при миеломной болезни.
Мочевина играет важную роль в механизмах концентрирования мочи, свободно фильтруется в клубочках. В проксимальном канальце часть мочевины пассивно реабсорбируется за счет градиента концентрации, который возникает вследствие концентрирования мочи. Остальная часть мочевины доходит до собирательных трубочек. В собирательных трубочках под влиянием АДГ происходит реабсорбция воды и концентрация мочевины повышается. АДГ усиливает проницаемость стенки и для мочевины, и она переходит в мозговое вещество почки, создавая здесь примерно 50% осмотического давления. Из интерстиция по концентрационному градиенту мочевина диффундирует в петлю Генле и вновь поступает в дистальные канальцы и собирательные трубочки. Таким образом, совершается внутрипочечный круговорот мочевины. В случае водного диуреза всасывание воды в дистальном отделе нефрона прекращается, а мочевины выводится больше. Таким образом, ее экскреция зависит от диуреза.
Реабсорбция слабых кислот и оснований зависит от того, в какой форме они находятся - в ионизированной или неионизированной. Слабые основания и кислоты в ионизированном состоянии не реабсорбируются и выводятся с мочой. Степень ионизации оснований увеличивается в кислой среде, поэтому они с большей скоростью экскретируются с кислой мочой, слабые кислоты, напротив, быстрее выводятся с щелочной мочой. Это имеет большое значение, так как многие лекарственные вещества являются слабыми основаниями или слабыми кислотами. Поэтому при отравлении ацетилсалициловой кислотой или фенобарбиталом (слабыми кислотами) необходимо вводить щелочные растворы (NaHCO3), для того чтобы перевести эти кислоты в ионизированное состояние, тем самым способствуя их быстрому выведению из организма. Для быстрой экскреции слабых оснований необходимо вводить в кровь кислые продукты для закисления мочи.
Вода реабсорбируется во всех отделах нефрона пассивно за счет транспорта осмотически активных веществ: глюкозы, аминокислот, белков, ионов натрия, калия, кальция, хлора. При снижении реабсорбции осмотически активных веществ уменьшается и реабсорбция воды. Наличие глюкозы в конечной моче ведет к увеличению диуреза (полиурии).
Основным ионом, обеспечивающим пассивное всасывание воды, является натрий. Натрий, как указывалось выше, также необходим для транспорта глюкозы и аминокислот. Кроме того, он играет важную роль в создании осмотически активной среды в интерстиции мозгового слоя почки, благодаря чему происходит концентрирование мочи.
Поступление натрия из первичной мочи через апикальную мембрану внутрь клетки канальцевого эпителия происходит пассивно по электрохимическому и концентрационному градиентам. Выведение натрия из клетки через базолатеральные мембраны осуществляется активно с помощью Na+/K+-АТФазы. Так как энергия клеточного метаболизма расходуется на перенос натрия, транспорт его является первично-активным. Транспорт натрия в клетку может происходить за счет разных механизмов. Один из них - это обмен Na+ на Н+ (противоточный транспорт, или антипорт). В этом случае ион натрия переносится внутрь клетки, а ион водорода - наружу. Другой путь переноса натрия в клетку осуществляется с участием аминокислот, глюкозы. Это так называемый котранспорт, или симпорт. Частично реабсорбция натрия связана с секрецией калия.
Сердечные гликозиды (строфантин К, оубаин) способны угнетать фермент Na+/К+-АТФазу, обеспечивающую перенос натрия из клетки в кровь и транспорт калия из крови в клетку.
Большое значение в механизмах реабсорбции воды и ионов натрия, а также концентрирования мочи имеет работа так называемой поворотно-противоточной множительной системы. После прохождения проксимального отрезка канальца изотоничный фильтрат в уменьшенном объеме поступает в петлю Генле. В этом участке интенсивная реабсорбция натрия не сопровождается реабсорбцией воды, так как стенки этого отрезка мало проницаемы для воды даже под воздействием АДГ. В связи с этим наступают разведение мочи в просвете нефрона и концентрация натрия в интерстиции. Разведенная моча в дистальном отделе канальца теряет избыток жидкости, становясь изотоничной плазме. Уменьшенный объем изотоничной мочи поступает в собирательную систему, идущую в мозговом слое, высокое осмотическое давление в интерстиции которого обусловлено повышенной концентрацией натрия. В собирательных трубочках под влиянием АДГ продолжается обратное всасывание воды в соответствии с концентрационным градиентом. Проходящие в мозговом слое vasa recta функционируют как противоточно-обменные сосуды, забирающие по пути к сосочкам натрий и отдающие его до возвращения к корковому слою. В глубине мозгового слоя таким путем поддерживается высокое содержание натрия, обеспечивающее резорбцию воды из собирательной системы и концентрацию мочи.
Функциями канальцевого аппарата почки (включающего в себя проксимальный каналец, петлю нефрона, дистальный каналец и собирательные трубочки) являются:
— реабсорбция части профильтровавшихся в клубочке органических и неорганических веществ;
— секреция в просвет канальца веществ, содержащихся в крови или образующихся в клетках канальцев,
— концентрирование мочи.
Реабсорбция – это обратное всасывание различных веществ из просвета канальцев в плазму перитубулярных капилляров. Реабсорбция происходит во всех отделах канальцев нефрона, в собирательной трубочке и определяется особенностями строения канальцевого эпителия почек. Поверхность клеток проксимального извитого канальца, обращенная в его просвет имеет покрытую гликокаликсом густую щеточную каемку, которая в 40 раз увеличивает площадь контакта мембраны с канальцевой жидкостью. Под щеточной каемкой между клетками имеются проницаемые плотные соединения.
Апикальную часть плазмолеммы называют также люминальной, она обладает высокой ионной проницаемостью, содержит различные белки-переносчики и обеспечивает преимущественно пассивный транспорт различных веществ.
Базолатеральная часть клетки увеличена за счет складчатости мембраны и содержит большое число митохондрий, что определяет сосредоточенность в ней систем активного транспорта (ионных насосов).
Пороговая реабсорбция отражает зависимость всасывания вещества от его концентрации в плазме крови. Если концентрация вещества в плазме не превышает определенный пороговый уровень, то это вещество будет полностью реабсорбировано в канальцах нефрона, если же превышает – то реабсорбируется не полностью и появляется в конечной моче, что связано с максимальным насыщением переносчиков.
Первичная моча,
проходя по канальцах и уборочных трубочках, перед тем как превратиться в конечную мочу, претерпевает значительные изменения. Разница состоит не только в ее количестве (с 180 л остается 1-1,5 л), но и качества. Некоторые вещества, нужные организму, полностью исчезают из мочи или их становится гораздо меньше. Происходит процесс реабсорбции
. Концентрация других веществ во много раз увеличивается: они концентрируются при реабсорбции воды. Еще другие вещества, которых вообще не было в первичной мочи,
появляются в конечной. Это происходит в результате их секреции.
Процессы реабсорбции могут быть активными или пассивными. Для осуществления активного процесса необходимо, чтобы были специфические транспортные системы и энергия. Пассивные процессы происходят, как правило, без затраты энергии по законам физики и химии.
Канальцевая реабсорбция происходит во всех отделах, но ее механизм в разных частях неодинакова. Условно можно выделить С отделы : проксимальный извитой каналец, петля нефрона и дистальный извитой каналец С уборочной трубочкой.
В проксимальных извитых канальцах полностью реабсорбируются аминокислоты, глюкоза, витамины, белки, микроэлементы. В этом же отделе реабсорбируется около 2/3 воды и неорганических солей Na +, К + Са2 +, Mg2 +, Cl-, НС07, т.е. вещества, которые нужны организму для его деятельности. Механизм реабсорбции главным образом прямо или косвенно связан с реабсорбцией Na+.
Реабсорбция натрия.
Большая часть Na + реабсорбируется против градиента концентрации за счет энергии АТФ. Реабсорбция Na + осуществляется в 3 этапа:
перенос иона через апикальную мембрану эпителиальных клеток канальцев, транспортировки в базальной или латеральной мембраны и перенос через указанные мембраны в межклеточную жидкость и в кровь. Основной движущей силой реабсорбции является перенос Na + с помощью Na +, К +-АТФ-азы через базолатерального мембрану. Это обеспечивает постоянное отток ионов. Вследствие этого Na + по градиенту концентрации с помощью специальных образований эндоплазматического ретикулума поступает к мембранам, возвращенных в межклеточной среды. Вследствие этого постоянно действующего конвейера концентрация ионов внутри клетки и особенно вблизи апикальной мембраны становится гораздо ниже, чем с другой ее стороны, это способствует пассивному поступлению Na+ в клетку по ионному градиенту. Таким образом,
2 этапа натриевой реабсорбции клетками канальцев являются пассивными и только один, конечный, требует затрат энергии. Кроме того, часть Na + реабсорбируется пассивно по межклеточных промежутках вместе с водой.
Глюкоза. Глюкоза реабсорбируется вместе с транспортом Na+. В апикальной мембране клеток есть специальные транспортеры. Это белки с молекулярной массой 320 000, которые в начальных отделах проксимального канальца переносят друг Na + и одну молекулу глюкозы (постепенное уменьшение концентрации глюкозы в моче приводит к тому, что в следующей области канальца для переноса одной молекулы глюкозы используется уже два Na +). Движущей силой этого процесса является также электрохимический градиент Na+. На противоположной стороне клетки комплекс Na — глюкоза — переносчик распадается на три элемента . Вследствие этого освобожден переносчик возвращается на свое прежнее место и снова приобретает способность переносить новые комплексы Na + и глюкозы. В клетке концентрация глюкозы увеличивается, благодаря чему образуется градиент концентрации, который направляет его в базально-латеральных мембран клетки и обеспечивает выход в межклеточную жидкость. Отсюда глюкоза поступает в кровеносные капилляры и возвращается в общий кровоток. Апикальная мембрана не пропускает глюкозу обратно в просвет канальца. Транспортные переносчики глюкозы содержатся лишь в проксимальном отделе канальцев, поэтому глюкоза реабсорбируется только здесь.
В норме при обычном уровне глюкозы в крови, а следовательно и концентрации ее в первичной мочи, реабсорбируется вся глюкоза. Однако при повышении уровня глюкозы в крови более 10 ммоль/л (около 1,8 г/л) мощность транспортных систем становится недостаточной для реабсорбции. Первые следы нереабсорбованои глюкозы в конечной моче обнаруживаются при превышении его концентрации в крови. Чем выше концентрация глюкозы в крови, тем большее количество нереабсорбованои глюкозы. До концентрации ее 3,5 г/л это увеличение еще не прямо пропорционально, поскольку в процесс еще не включается часть транспортеров. Но, начиная с уровня 3,5 г/л , выведение глюкозы с мочой становится прямо пропорционален концентрации ее в крови. У мужчин полная нагрузка системы реабсорбции наблюдается при поступлении 2,08 ммоль/мин (375 мг/мин) глюкозы, а у женщин — 1, 68 ммоль/мин (303 мг/мин) из расчета на 1,73 м2 поверхности тела.
Аминокислоты. Реабсорбция аминокислот происходит по такому же механизму, как и реабсорбция глюкозы. Полная реабсорбция аминокислот происходит уже в начальных отделах проксимальных канальцев. Этот процесс связан с активной реабсорбцией Na + через апикальную мембрану клеток. Выявлено 4 типа транспортных систем: а) для основных б) для кислых в) для гидрофильных г) для гидрофобных аминокислот. С клетки аминокислоты пассивно по градиенту концентрации проходят через базальную мембрану в межклеточную жидкость, а оттуда — в кровь. Появление аминокислот в моче может быть следствием нарушения транспортных систем или очень высокой концентрации его в крови. В последнем случае может проявляться эффект, который по механизму напоминает глюкозурию — перегрузка транспортных систем. Иногда наблюдается конкуренция кислот одного типа за общий переносчик.
Белки. Механизм реабсорбции белков значительно отличается от механизма реабсорбции описанных соединений. Попадая в первичную мочу, небольшое количество белков в норме почти полностью реабсорбируется путем пиноцитоза. В цитоплазме клеток проксимальных канальцев белки распадаются при участии лизосомальных ферментов. Аминокислоты, которые образуются, по градиенту концентрации из клетки поступают в межклеточную жидкость, а оттуда — в кровеносные капилляры. Таким путем может реабсорбуватися до ЗО мг белка за 1 мин. При повреждении клубочков в фильтрат попадает больше белков и часть может поступать в мочу (протеинурия).
Канальцевая секреция. В современной физиологической литературе, касающейся деятельности почек, термин секреция имеет два значения. Первое из них описывает процесс переноса вещества через клетки из крови в просвет канальца в неизменном виде, что увеличивает скорость экскреции вещества почкой. Второе - выделение из клетки в кровь или в просвет канальца синтезированных в почке физиологически активных веществ (например, простагландины, брадикинин и др.) или экскретируемых веществ (например, гиппуровая кислота).
Секреция органических и неорганических веществ - один из важных процессов, обеспечивающих процесс мочеобразования. У рыб некоторых видов в почке отсутствуют клубочки. В таких случаях секреция играет ведущую роль в деятельности почки. В почках большинства других классов позвоночных, в том числе и у млекопитающих, секреция обеспечивает выделение из крови в просвет канальцев дополнительных количеств некоторых веществ, которые могут фильтроваться ив почечных клубочках.
Таким образом, секреция ускоряет выделение почкой некоторых чужеродных веществ, конечных продуктов обмена, ионов. В почке у млекопитающих секретируются органические кислоты (пенициллин, парааминогиппуровая кислота - ПАГ, диодраст, мочевая кислота), органические основания (холин, гуанидин), неорганические вещества (калий). Почка гломерулярных и агломерулярных морских костистых рыб способна к секреции ионов магния, кальция, сульфатов. Различаются места секреции разных веществ. В почке всех позвоночных местом секреции органических кислот и оснований служат клетки проксимального сегмента нефрона, особенно его прямой части, секреция калия преимущественно происходит в клетках дистального извитого канальца и собирательных трубок.
Механизм процесса секреции органических кислот. Рассмотрим этот процесс на примере выделения почкой ПАГ. После введения в кровь ПАГ ее секреция почкой нарастает и очищение от нее крови значительно превышает величину очищения крови от одновременно введенного инулина. Это означает, что ПАГ не только фильтруется в клубочках, но и помимо клубочков в просвет нефрона поступают значительные ее количества. Экспериментально было показано, что такой процесс обусловлен секрецией ПАГ из крови в просвет проксимальных отделов канальцев. В мембране клетки этого канальца, обращенной к межклеточной жидкости, имеется переносчик (котранспортер), обладающий высоким сродством к ПАГ. В присутствии ПАГ образуется комплекс переносчика с ПАГ, который перемещается в мембране и на ее внутренней поверхности распадается, высвобождая ПАГ в цитоплазму, а переносчик приобретает снова способность перемещаться к внешней поверхности мембраны и соединяться с новой молекулой ПАГ. Механизм секреции органических кислот включает ряд этапов. В базальной плазматической мембране имеется Na+, K+-АТФаза, которая удаляет из клетки ионы Na+ и способствует поступлению в клетку ионов К+. Более низкая концентрация в цитоплазме ионов Na+ позволяет поступать внутрь клетки ионам Na+ по градиенту концентрации при участии натриевых котранспортеров. Один из типов такого котранспортера способствует поступлению через базальную плазматическую мембрану α-кетоглутарата и Na+. В этой же мембране имеется анионный обменник, который удаляет из цитоплазмы α-кетоглутарат в обмен на поступающий из межклеточной жидкости в клетку парааминогиппурат (ПАГ), диодраст или некоторые иные органические кислоты. Это вещество движется по клетке в сторону люминальной мембраны и через нее проходит в просвет канальца по механизму облегченной диффузии.
Угнетение дыхания цианидами, разобщение дыхания и окислительного фосфорилирования динитрофенолом снижает и прекращает секрецию. В обычных физиологических условиях уровень секреции зависит от числа переносчиков в мембране. Секреция ПАГ возрастает пропорционально увеличению концентрации ПАГ в крови до тех пор, пока все молекулы переносчика не насытятся ПАГ. Максимальная скорость транспорта ПАГ достигается в тот момент, когда количество ПАГ, доступное для транспорта, равно количеству молекул переносчика, которые могут образовывать комплекс с ПАГ. Эта величина определяется как максимальная способность к транспорту ПАГ - Ттран. Поступившая в клетку ПАГ движется по цитоплазме к апикальной мембране и через нее специальным механизмом выделяется в просвет канальца.
Билет 15
Предыдущая3456789101112131415161718Следующая
ПОЧКИ И ИХ ФУНКЦИИ
Канальцевая реабсорбция
Начальный этап мочеобразования, приводящий к фильтрации всех низкомолекулярных компонентов плазмы крови, неизбежно должен сочетаться с существованием в почке систем, реабсорбирующих все ценные для организма вещества. В обычных условиях в почке человека за сутки образуется до 180 л фильтрата, а выделяется 1,0-1,5 л мочи, остальная жидкость всасывается в канальцах. Роль клеток различных сегментов нефрона в реабсорбции неодинакова. Проведенные на животных опыты с извлечением микропипеткой жидкости из различных участков нефрона позволили выяснить особенности реабсорбции различных веществ в разных частях почечных канальцев (рис. 12.6). В проксимальном сегменте нефрона практически полностью реабсорбируются аминокислоты, глюкоза, витамины, белки, микроэлементы, значительное количество ионов Na+, СI-,НСОз. В последующих от делах нефрона всасываются преимущественно электролиты и вода.
Реабсорбция натрия и хлора представляет собой наиболее значительный по объему и энергетическим тратам процесс. В проксимальном канальце в результате реабсорбции большинства профильтровавшихся веществ и воды объем первичной мочи уменьшается, и в начальный отдел петли нефрона поступает около ‘/з профильтровавшейся в клубочках жидкости. Из всего количества натрия, поступившего в нефрон при фильтрации, в петле нефрона всасывается до 25 %, в дистальном извитом канальце - около 9 %, и менее 1% реабсорбируется в собирательных трубках или экскретируется с мочой.
Реабсорбция в дистальном сегменте характеризуется тем, что клетки переносят меньшее, чем в проксимальном канальце, количество ионов, но против большего градиента концентрации. Этот сегмент нефрона и собирательные трубки играют важнейшую роль в регуляции объема выделяемой мочи и концентрации в ней осмотически активных веществ (осмотическая концентрация1). Б конечной моче концентрация натрия может снижаться до 1 ммоль/л по сравнению со 140 ммоль/л в плазме крови. В дистальном канальце калий не только реабсорбируется, но и секретируется при его избытке в организме.
В проксимальном отделе нефрона реабсорбция натрия, калия, хлора и других веществ происходит через высокопроницаемую для воды мембрану стенки канальца. Напротив, в толстом восходящем отделе петли нефрона, дистальных извитых канальцах и собирательных трубках реабсорбция ионов и воды происходит через малопроницаемую для воды стенку канальца; проницаемость мембраны для воды в отдельных участках нефрона и собирательных трубках может регулироваться, а.величина проницаемости изменяется в зависимости от функционального состояния организма (факультативная реабсорбция). Под влиянием импульсов, поступающих по эфферентным нервам, и при действии биологически активных веществ реабсорбция натрия и хлора регулируется в проксимальном отделе нефрона. Это особенно отчетливо проявляется в случае увеличения объема крови и внеклеточной жидкости, когда уменьшение реабсорбции в проксимальном канальце способствует усилению экскреции ионов и воды и тем самым - восстановлению водно-солевого равновесия. В проксимальном канальце всегда сохраняется изоосмия. Стенка канальца проницаема для воды, и объем реабсорбируемой воды определяется количеством реабсорбируемых осмотически активных веществ, за которыми вода движется по осмотическому градиенту. В конечных частях дистального сегмента нефрона и собирательных трубках проницаемость стенки канальца для воды регулируется вазопрессином.
Факультативная реабсорбция воды зависит от осмотической проницаемости канальцевой стенки, величины осмотического градиента и скорости движения жидкости по канальцу.
Для характеристики всасывания различных веществ в почечных канальцах существенное значение имеет представление о пороге выведения.
Непороговые вещества выделяются при любой их концентрации в плазме крови (и соответственно в ультрафильтрате). Такими веществами являются инулин, маннитол. Порог выведения практически всех физиологически важных, ценных для организма веществ различен. Так, выделение глюкозы с мочой (глюкозурия) наступает тогда, когда ее концентрация в клубочковом фильтрате (и в плазме крови) превышает 10 ммоль/л. Физиологический смысл этого явления будет раскрыт при описании механизма реабсорбции.
Механизмы канальцевой реабсорбции. Обратное всасывание различных веществ в канальцах обеспечивается активным и пассивным транспортом. Если вещество реабсорбируется против электрохимического и концентрационного градиентов, процесс называется активным транспортом. Различают два вида активного транспорта - первично-активный и вторично-активный. Первично-активным транспорт называется в том случае, когда происходит перенос вещества против электрохимического градиента за счет энергии клеточного метаболизма. Примером служит транспорт ионов Na+, который происходит при участии фермента Na+, К+-АТФазы, использующей энергию АТФ. Вторично-активным называется перенос вещества против концентрационного градиента, но без затраты энергии клетки непосредственно на этот процесс; так реабсорбируются глюкоза, аминокислоты. Из просвета канальца эти органические вещества поступают в клетки проксимального канальца с помощью специального переносчика, который обязательно должен присоединить ион Na+. Этот комплекс (переносчик + органическое вещество + Na+) способствует перемещению вещества через мембрану щеточной каемки и его поступлению внутрь клетки. Движущей силой переноса этих веществ через апикальную плазматическую мембрану служит меньшая по сравнению с просветом канальца концентрация натрия в цитоплазме клетки. Градиент концентрации натрия обусловлен непрестанным активным выведением натрия из клетки во внеклеточную жидкость с помощью Na+, К+-АТФазы, локализованной в латеральных и базальной мембранах клетки.
Реабсорбция воды, хлора и некоторых других ионов, мочевины осуществляется с помощью пассивного транспорта - по электрохимическому, концентрационному или осмотическому градиенту. Примером пассивного транспорта является реабсорбция в дистальном извитом канальце хлора по электрохимическому градиенту, создаваемому активным транспортом натрия. По осмотическому градиенту транспортируется вода, причем скорость ее всасывания зависит от осмотической проницаемости стенки канальца и разности концентрации осмотически активных веществ по обеим сторонам его стенки. В содержимом проксимального канальца вследствие всасывания воды и растворенных в ней веществ растет концентрация мочевины, небольшое количество которой по концентрационному градиенту реабсорбируется в кровь.
Достижения в области молекулярной биологии позволили установить строение молекул ионных и водных каналов (аквапоринов) рецепторов, аутакоидов и гормонов и тем самым проникнуть в сущность некоторых клеточных механизмов, обеспечивающих транспорт веществ через стенку канальца. Различны свойства клеток разных отделов нефрона, неодинаковы свойства цитоплазматической мембраны в одной и той же клетке. Апикальная мембрана клетки, обращенная в просвет канальца, имеет иные характеристики, чем ее базальная и боковые мембраны, омываемые межклеточной жидкостью и соприкасающиеся с кровеносным капилляром. Вследствие этого апикальная и базальная плазматические мембраны участвуют в транспорте веществ по-разному; специфично и действие биологически активных веществ на ту и другую мембраны.
Клеточный механизм реабсорбции ионов рассмотрим на примере Na+. В проксимальном канальце нефрона всасывание Na+ в кровь происходит в результате ряда процессов, один из которых - активный транспорт Na+ из просвета канальца, другой - пассивная реабсорбция Na+ вслед за активно транспортируемыми в кровь как ионами гидрокарбоната, так и С1-. При введении одного микроэлектрода в просвет канальцев, а второго - в околоканальцевую жидкость было выявлено, что разность потенциалов между наружной и внутренней поверхностью стенки проксимального канальца оказалась очень небольшой - около 1,3 мВ, в области дистального канальца она может достигать- 60 мВ (рис.12.7). Просвет обоих канальцев электроотрицателен, а в крови (следовательно, и во внеклеточной жидкости), концентрация Na+ выше, чем в жидкости, находящейся в просвете этих канальцев, поэтому реабсорбция Na+ осуществляется активно против градиента электрохимического потенциала. При этом из просвета канальца Na+ входит в клетку по натриевому каналу или при участии переносчика. Внутренняя часть клетки запряжена отрицательно, и положительно заряженный Na+ поступает в клетку по градиенту потенциала, движется в сторону базальной плазматической мембраны, через которую натриевым насосом выбрасывается в межклеточную жидкость; градиент потенциала на этой мембране достигает 70-90 мВ.
Имеются вещества, которые могут влиять на отдельные элементы системы реабсорбции Na+. Так, натриевый канал в мембране клетки дистального канальца и собирательной трубки блокируется амилоридом и триамтереном, в результате чего Na+ не может войти в канал. Б клетках имеется несколько типов ионных насосов.
Канальцевая реабсорбция и ее регуляция
Один из них представляет собой Na+, К+-АТФазу. Этот фермент находится в базальной и латеральных мембранах клетки и обеспечивает транспорт Na+ из клетки в кровь и поступление из крови в клетку К+. Фермент угнетается сердечными гликозидами, например строфантином, уабаином. В реабсорбции гидрокарбоната важная роль принадлежит ферменту карбоангидразе, ингибитором которого является ацетазоламид -он прекращает реабсорбцию гидрокарбоната, который экскретируется с мочой.
Фильтруемая глюкоза практически полностью реабсорбируется клетками проксимального канальца, и в норме за сутки с мочой выделяется незначительное ее количество (не более 130 мг). Процесс обратного всасывания глюкозы осуществляется против высокого концентрационного градиента и является вторично-активным. В апикальной (люминальной) мембране клетки глюкоза соединяется с переносчиком, который должен присоединить также Na+, после чего комплекс транспортируется через апикальную мембрану, т. е. в цитоплазму поступают глюкоза и Na+. Апикальная мембрана отличается высокой селективностью и односторонней проницаемостью и не пропускает ни глюкозу, ни Na+ обратно из клетки в просвет канальца. Эти вещества движутся к основанию клетки по градиенту концентрации. Перенос глюкозы из клетки в кровь через базальную плазматическую мембрану носит характер облегченной диффузии, a Na+, как уже отмечалось выше, удаляется натриевым насосом, находящимся в этой мембране.
Аминокислоты почти полностью реабсорбируются клетками проксимального канальца. Имеется не менее 4 систем транспорта аминокислот из просвета канальца в кровь, осуществляющих реабсорбцию нейтральных, двуосновных, дикарбоксильных аминокислот и иминокислот. Каждая из этих систем обеспечивает всасывание ряда аминокислот одной группы. Так, система реабсорбции двуосновных аминокислот участвует во всасывании лизина, аргинина, орнитина и, возможно, цистина. При введении в кровь избытка одной из этих аминокислот начинается усиленная экскреция почкой аминокислот только данной группы. Системы транспорта отдельных групп аминокислот контролируются раздельными генетическими механизмами. Описаны наследственные заболевания, одним из проявлений которых служит увеличенная экскреция определенных групп аминокислот (аминоацидурия).
Выделение с мочой слабых кислот и оснований зависит от их клубочковой фильтрации, процесса реабсорбции или секреции. Процесс выведения этих веществ во многом определяется «неионной диффузией», влияние которой особенно сказывается в дистальных канальцах и собирательных трубках. Слабые кислоты и основания могут существовать в зависимости от рН среды в двух формах - неионизированной и ионизированной. Клеточные мембраны более проницаемы для неионизированных веществ. Многие слабые кислоты с большей скоростью экскретируются с щелочной мочой, а слабые основания, напротив, - с кислой. Степень ионизации оснований увеличивается в кислой среде, но уменьшается в щелочной. В неионизированном состоянии эти вещества через липиды мембран проникают в клетки, а затем в плазму крови, т. е. они реабсорбируются. Если значение рН канальцевой жидкости сдвинуто в кислую сторону, то основания ионизируются, плохо всасываются и экскретируются с мочой. Никотин - слабое основание, при рН 8,1 ионизируется 50 %, в 3-4 раза быстрее экскретируется с кислой (рН около 5), чем с щелочной (рН 7,8) мочой. Процесс «неионной диффузии» влияет на выделение почками слабых оснований и кислот, барбитуратов и других лекарственных веществ.
Небольшое количество профильтровавшегося в клубочках белка реабсорбируется клетками проксимальных канальцев. Выделение белков с мочой в норме составляет не более 20-75 мг в сутки, а при заболеваниях почек оно может возрастать до 50 г в сутки. Увеличение выделения белков с мочой (протеинурия) может быть обусловлено нарушением их реабсорбции либо увеличением фильтрации.
В отличие от реабсорбции электролитов, глюкозы и аминокислот, которые, проникнув через апикальную мембрану, в неизмененном виде достигают базальной плазматической мембраны и транспортируются в кровь, реабсорбция белка обеспечивается принципиально иным механизмом. Белок попадает в клетку с помощью пиноцитоза. Молекулы профильтровавшегося белка адсорбируются на поверхности апикальной мембраны клетки, при этом мембрана участвует в образовании пиноцитозной вакуоли. Эта вакуоль движется в сторону базальной части клетки. В околоядерной области, где локализован пластинчатый комплекс (аппарат Гольджи), вакуоли могут сливаться с лизосомами, обладающими высокой активностью ряда ферментов. В лизосомах захваченные белки расщепляются и образовавшиеся аминокислоты, дипептиды удаляются в кровь через базальную плазматическую мембрану. Следует, однако, подчеркнуть, что не все белки подвергаются гидролизу в процессе транспорта и часть их переносится в кровь в неизмененном виде.
Определение величины реабсорбции в канальцах почки. Обратное всасывание веществ, или, иными словами, их транспорт (Т) из просвета канальцев в тканевую (межклеточную) жидкость и в кровь, при реабсорбции R (TRX) определяется по разности между количеством вещества X (F∙Px∙fx), профильтровавшегося в клубочках, и количеством вещества, выделенного с мочой (UX ∙V).
TRX =F∙px.fx ─Ux∙V,
где F - объем клубочковой фильтрации, fx - фракция вещества X, не связанная с белками в плазме по отношению к его об щей концентрации в плазме крови, Р - концентрация вещества в плазме крови, U - концентрация вещества в моче.
По приведенной формуле рассчитывают абсолютное количество реабсорбируемого вещества. При вычислении относительной реаб-сорбции (% R) определяют долю вещества, подвергшуюся обратному всасыванию по отношению к количеству вещества, профильтровавшегося в клубочках:
% R= (1 - EFX)∙100.
Для оценки реабсорбционной способности клеток проксимальных канальцев важное значение имеет определение максимальной величины транспорта глюкозы (TmG). Эту величину измеряют при полном насыщении глюкозой системы ее канальцевого транспорта (см. рис. 12.5). Для этого вливают в кровь раствор глюкозы и тем самым повышают ее концентрацию в клубочковом фильтрате до тех пор, пока значительное количество глюкозы не начнет выделяться с мочой:
TmG=F∙PG-UG∙ V,
где F - клубочковая фильтрация, РG - концентрация глюкозы в плазме крови, a UG - концентрация глюкозы в моче; Тт - максимальный канальцевый транспорт изучаемого вещества. Величина ТmG характеризует полную загрузку системы транспорта глюкозы; у мужчин эта величина равна 375 мг/мин, а у женщин - 303 мг/мин при расчете на 1,73 м2 поверхности тела.
Канальцевая реабсорбция
Первичная моча превращается в конечную благодаря процессам, которые происходят в почечных канальцах и собирательных бочках. В почке человека за сутки образуется 150 — 180 л фильма, или первичной мочи, а выделяется 1,0-1,5 л мочи. Остальная жидкость всасывается в канальцах и собирательных трубочках.
Канальцевая реабсорбция — это процесс обратного всасывания воды и веществ из содержащейся в просвете канальцев мочи в лимфу и кровь. Основной смысл реабсорбции состоит в том, чтобы сохранить организму все жизненно важные вещества в необходимых количествах. Обратное всасывание происходит во всех отделах нефрона. Основная масса молекул реабсорбируется в проксимальном отделе нефрона. Здесь практически полностью абсорбируются аминокислоты, глюкоза, витамины, белки, микроэлементы, значительное количество ионов Na+, C1-, HCO3- и многие другие вещества.
Схема канальцевой реабсорбации
В петле Генле, дистальном отделе канальца и собирательных трубочках всасываются электролиты и вода. Ранее считали, что реабсорбция в проксимальной части канальца является обязательной и нерегулируемой. В настоящее время доказано, что она регулируется как нервными, так и гуморальными факторами.
Обратное всасывание различных веществ в канальцах может происходить пассивно и активно. Пассивный транспорт происходит без затраты энергии по электрохимическому, концентрационному или осмотическому градиентам. С помощью пассивного транспорта осуществляется реабсорбция воды, хлора, мочевины.
Активным транспортом называют перенос веществ против электрохимического и концентрационного градиентов. Причем различают первично-активный и вторично-активный транспорт. Первично-активный транспорт происходит с затратой энергии клетки. Примером служит перенос ионов Na+ с помощью фермента Na+, K+ — АТФазы, использующей энергию АТФ. При вторично-активном транспорте перенос вещества осуществляется за счет энергии транспорта другого вещества. Механизмом вторично-активного транспорта реабсорбируются глюкоза и аминокислоты.
Глюкоза. Она поступает из просвета канальца в клетки проксимального канальца с помощью специального переносчика, который должен обязательно присоединить ион Ма4′. Перемещение этого комплекса внутрь клетки осуществляется пассивно по электрохимическому и концентрационному градиентам для ионов Na+. Низкая концентрация натрия в клетке, создающая градиент его концентрации между наружной и внутриклеточной средой, обеспечивается работой натрий-калиевого насоса базальной мембраны.
В клетке этот комплекс распадается на составные компоненты. Внутри почечного эпителия создается высокая концентрация глюкозы, поэтому в дальнейшем по градиенту концентрации глюкоза переходит в интерстициальную ткань. Этот процесс осуществляется с участием переносчика за счет облегченной диффузии. Далее глюкоза уходит в кровоток. В норме при обычной концентрации глюкозы в крови и, соответственно, в первичной моче вся глюкоза реабсорбируется. При избытке глюкозы в крови, а значит, в первичной моче может произойти максимальная загрузка канальцевых систем транспорта, т.е. всех молекул-переносчиков.
В этом случае глюкоза больше не сможет реабсорбироваться и появится в конечной моче (глюкозурия). Эта ситуация характеризуется понятием " максимальный канальцевый транспорт" (Тм). Величине максимального канальцевого транспорта соответствует старое понятие " почечный порог выведения". Для глюкозы эта величина составляет 10 ммоль/л.
Вещества, реабсорбция которых не зависит от их концентрации в плазме крови, называются непороговыми. К ним относятся вещества, которые или вообще не реабсорбируются, (инулин, маннитол) или мало реабсорбируются и выделяются с мочой пропорционально накоплению их в крови (сульфаты).
Аминокислоты. Реабсорбция аминокислот происходит также по механизму сопряженного с Na+ транспорта. Профильтровавшиеся в клубочках аминокислоты на 90% реабсорбируются клетками проксимального канальца почки. Этот процесс осуществляется с помощью вторично-активного транспорта, т.е. энергия идет на работу натриевого насоса. Выделяют не менее 4 транспортных систем для переноса различных аминокислот (нейтральных, двуосновных, дикарбоксильных и аминокислот). Эти же системы транспорта действуют и в кишечнике для всасывания аминокислот.
Канальцевая реабсорбция
Описаны генетические дефекты, когда определенные аминокислоты не реабсорбируются и не всасываются в кишечнике.
Белок. В норме небольшое количество белка попадает в фильтрат и реабсорбируется. Процесс реабсорбции белка осуществляется с помощью пиноцитоза. Эпителий почечного канальца активно захватывает белок. Войдя в клетку, белок подвергается гидролизу со стороны ферментов лизосом и превращается в аминокислоты. Не все белки подвергаются гидролизу, часть их переходит в кровь в неизмененном виде. Этот процесс активный и требует энергии. За сутки с конечной мочой уходит не более 20-75 мг белка. Появление белка в моче носит название протеинурии. Протеинурия может быть и в физиологических условиях, пример, после тяжелой мышечной работы. В основном протеинурия имеет место в патологии при нефритах, нефропатиях, при миеломной болезни.
Мочевина. Она играет важную роль в механизмах концентрирования мочи, свободно фильтруется в клубочках. В проксимальном канальце часть мочевины пассивно реабсорбируется за счет градиента концентрации, который возникает вследствие концентрирования мочи. Остальная часть мочевины доходит до собирательных трубочек. В собирательных трубочках под влиянием АДГ происходит реабсорбция воды и концентрация мочевины повышается. АДГ усиливает проницаемость стенки и для мочевины, и она переходит в мозговое вещество почки, создавая здесь примерно 50% осмотического давления.
Из интерстиция по концентрационному градиенту мочевина диффундирует в петлю Генле и вновь поступает в дистальные канальцы и собирательные трубочки. Таким образом совершается внутрипочечный круговорот мочевины. В случае водного диуреза всасывание воды в дистальном отделе нефрона прекращается, а мочевины выводится больше. Таким образом ее экскреция зависит от диуреза.
Слабые органические кислоты и основания. Реабсорбция слабых кислот и оснований зависит от того, в какой форме они находятся — в ионизированной или неионизированной. Слабые основания и кислоты в ионизированном состоянии не реабсорбируются и выводятся с мочой. Степень ионизации оснований увеличивается в кислой среде, поэтому они с большей скоростью экскретируются с кислой мочой, слабые кислоты, напротив, быстрее выводятся с щелочной мочой.
Это имеет большое значение, так как многие лекарственные вещества являются слабыми основаниями или слабыми кислотами. Поэтому при отравлении ацетилсалициловой кислотой или фенобарбиталом (слабыми кислотами) необходимо вводить щелочные растворы (NaHCO3), для того чтобы перевести эти кислоты в ионизированное состояние, тем самым способствуя их быстрому выведению из организма. Для быстрой экскреции слабых оснований необходимо вводить в кровь кислые продукты для закисления мочи.
Вода и электролиты. Вода реабсорбируется во всех отделах нефрона. В проксимальных извитых канальцах реабсорбируется около 2/3 всей воды. Около 15% реабсорбируется в петле Генле и 15% — в дистальных извитых канальцах и собирательных трубочках. Вода реабсорбируется пассивно за счет транспорта осмотически активных веществ: глюкозы, аминокислот, белков, ионов натрия, калия, кальция, хлора. При снижении реабсорбции осмотически активных веществ уменьшается и реабсорбция воды. Наличие глюкозы в конечной моче ведет к увеличению диуреза (полиурии).
Основным ионом, обеспечивающим пассивное всасывание воды, является натрий. Натрий, как указывалось выше, также необходим для транспорта глюкозы и аминокислот. Кроме Того, он играет важную роль в создании осмотически активной среды в интерстиции мозгового слоя почки, благодаря чему происходит концентрирование мочи. Реабсорбция натрия совершается во всех отделах нефрона. Около 65% ионов натрия реабсорбируется в проксимальных канальцах, 25% — в петле нефрона, 9% — в дистальном извитом канальце и 1% — в собирательных трубочках.
Поступление натрия из первичной мочи через апикальную мембрану внутрь клетки канальцевого эпителия происходит пассивно по электрохимическому и концентрационному градиентам. Выведение натрия из клетки через базолатеральные мембраны осуществляется активно с помощью Na+, K+ — АТФазы. Так как энергия клеточного метаболизма расходуется на перенос натрия, транспорт его является первично-активным. Транспорт натрия в клетку может происходить за счет разных механизмов. Один из них — это обмен Na+ на Н+ (противоточный транспорт, или антипорт). В этом случае ион натрия переносится внутрь клетки, а ион водорода — наружу.
Другой путь переноса натрия в клетку осуществляется с участием аминокислот, глюкозы. Это так называемый котранспорт, или симпорт. Частично реабсорбция натрия связана с секрецией калия.
Сердечные гликозиды (строфантин К, оубаин) способны угнетать фермент Na+, К+ — АТФазу, обеспечивающую перенос натрия из клетки в кровь и транспорт калия из крови в клетку.
Большое значение в механизмах реабсорбции воды и ионов натрия, а также концентрирования мочи имеет работа так называемой поворотно-противоточной множительной системы.
Поворотно-противоточная система представлена параллельно расположенными коленами петли Генле и собирательной трубочкой, по которым жидкость движется в разных направлениях (противоточно). Эпителий нисходящего отдела петли пропускает воду, а эпителий восходящего колена непроницаем для воды, но способен активно переносить ионы натрия в тканевую жидкость, а через нее обратно в кровь. В проксимальном отделе происходит всасывание натрия и воды в эквивалентных количествах и моча здесь изотонична плазме крови.
В нисходящем отделе петли нефрона реабсорбируется вода и моча становится более концентрированной (гипертонической). Отдача воды происходит пассивно за счет того, что в восходящем отделе одновременно осуществляется активная реабсорбция ионов натрия. Поступая в тканевую жидкость, ионы натрия повышают в ней осмотическое давление, тем самым способствуя притягиванию в тканевую жидкость воды из нисходящего отдела. В то же время повышение концентрации мочи в петле нефрона за счет реабсорбции воды облегчает переход натрия из мочи в тканевую жидкость. Так как в восходящем отделе петли Генле реабсорбируется натрий, моча становится гипотоничной.
Поступая далее в собирательные трубочки, представляющие собой третье колено противоточной системы, моча может сильно концентрироваться, если действует АДГ, повышающий проницаемость стенок для воды. В данном случае по мере продвижения по собирательным трубочкам в глубь мозгового вещества все больше и больше воды выходит в межтканевую жидкость, осмотическое давление которой повышено вследствие содержания в ней большого количества Na" 1" и мочевины, и моча становится все более концентрированной.
При поступлении больших количеств воды в организм почки, наоборот, выделяют большие объемы гипотонической мочи.
Канальцевая реабсорбция и секреция веществ в нефроне.
КАНАЛЬЦЕВАЯ РЕАБСОРБЦИЯ или обратное всасывание в кровь, содержащихся в первичной моче, воды, солей, органических веществ (глюкозы, белка, аминокислот, витаминов).
Результатом является уменьшение первичной мочи (на 70%), полное обратное всасывание в кровь полезных для метаболизма веществ (аминокислот, глюкозы, многих витаминов), частичное всасывание воды и ионов Na, Cl, K, Ca, выделение из крови в мочу токсических продуктов метаболизма (мочевины, мочевой кислоты, аммиака, креатинина, сульфатов, фосфатов).
Всасывание основных веществ осуществляется при помощи механизмов активного транспорта, диффузии и облегченной диффузии.
Например:
Главный ион, определяющий осмотическое давление, и, следовательно, реабсорбцию воды, Na+ входит в эпителиальные клетки пассивно, по градиенту концентрации, а затем выбрасывается с другой стороны клетки Na+-К+-АТФ-азой.
Ионы К+ реабсорбируются активно на апикальной мембране и затем выходят в кровь за счет диффузии.
В проксимальных извитых канальцах реабсорбируется 70% воды и ионов.
Реабсорбция катионов (Na+, K+, Ca2+, Mg2+) происходит против градиента концентраций, активно (с использованием энергии АТФ).
Отрицательно заряженные анионы притягиваются положительно заряженными катионами, и за счет электростатических сил поступают из мочи в кровь пассивно (Cl- и HCO3- вслед за Na+ и K+; SO42- и PO42- за Ca2+ и Mg2+), вода всасывается пассивно вслед за ионами по осмотическому градиенту.
Механизмы реабсорбции Ca2+, Mg2+, SO4-, PO4- сходны с механизмами реабсорбции Na+, K+ и Cl-.
Вещества могут переносятся в цитоплазму почечной эпителиальной клетки переносчиками совместно с ионами Na+.
При этом из эпителиальной клетки в кровь они поступают с помощью диффузии по градиенту концентрации.
При определенной концентрации веществ крови (порог выведения) эти вещества (пороговые) не будут полностью реабсорбироваться, и часть профильтровавшихся веществ окажется в конечной моче.
К пороговым веществам относится глюкоза, которая в норме (4,6-7,2 ммоль/л в крови) фильтруется, а затем полностью реабсорбируется.
При увеличении ее концентрации в крови до 10,8 ммоль/л часть глюкозы не будет успевать реабсорбироваться.
Она выделяется с мочой из организма и возникает глюкозурия.
РЕАБСОРБЦИЯ в различных участках нефрона неодинакова.
В ПРОКСИМАЛЬНОМ ОТДЕЛЕ реабсорбируются 40-45 % воды, натрия, бикарбонаты, хлор, аминокислоты, глюкоза, витамины, белки, микроэлементы к концу отдела — остается 1/3 ультрафильтрата с таким же осмотическим давлением как в плазме.
В ПЕТЛЕ ГЕНЛЕ реабсорбируется 25-28% воды, до 25% натрия, а также ионы хлора, калия, кальция, магния
В ДИСТАЛЬНОМ ОТДЕЛЕ — 10% воды, около 9% натрия, калия.
В СОБИРАТЕЛЬНЫХ ТРУБОЧКАХ — 20% воды, менее 1% натрия.
КАНАЛЬЦЕВАЯ СЕКРЕЦИЯ проявляется ВЫДЕЛЕНИЕМ из крови в ПРОСВЕТ КАНАЛЬЦЕВ продуктов обмена и чужеродных веществ
Канальцевая секреция является результатом активной деятельности ЭПИТЕЛИЯ почечных канальцев.
Она осуществляется против концентрационного или электрохимического градиента и позволяет быстро экскретировать органические основания и ионы, ЭПИТЕЛИАЛЬНЫЕ КЛЕТКИ секретируют из КРОВИ Холин, парааминогиппуровую кислоту, Видоизмененные молекулы лекарственных веществ и поглощают из ПЕРВИЧНОЙ МОЧИ Глютамин.
С помощью фермента глютаминазы РАСЩЕПЛЯЮТ глютамин на ГЛЮТАМИНОВУЮ КИСЛОТУ и АММИАК.
АММИАК выделяется в мочу, который выносится из организма в виде АММОНИЙНЫХ СОЛЕЙ.
Там же расщепляется Угольная кислота ферментом КАРБОАНГИДРАЗОЙ.
Как проходит процесс реабсорбции в почках
Ионы HСО3- всасываются в кровь (за счет электростатического притяжения их Na+ и К+).
Ионы H+ секретируются в мочу, с которой удаляются.
Этим объясняется кислая реакция конечной мочи (pH=4,5-6,5).
Этот механизм ПРЕДОХРАНЯЕТ организм от ЗАКИСЛЕНИЯ.
ЛОКАЛИЗАЦИЯ СЕКРЕЦИИ ВЕЩЕСТВ В НЕФРОНЕ различна
В ПРОКСИМАЛЬНОМ ОТДЕЛЕ секретируются Ионы водорода и Аммиак. Причем в извитой части секретируются органические основания:
Холин, Серотонин, Допамин, Хинин, морфин.
В прямой части – органические кислоты: парааминогиппуровая, Диодраст, Пенициллин, Мочевая кислота.
В ДИСТАЛЬНОМ ОТДЕЛЕ – парааминогиппуровая кислота, Аммиак, Ионы H+ и К+.
ЛЕКАРСТВЕННЫЕ ВЕЩЕСТВА выводятся из организма с помощью КЛУБОЧКОВОЙ ФИЛЬТРАЦИИ (левомицетин, стрептомицин, тетрациклин, неомицин, канамицин и др. антибиотики).
С помощью КАНАЛЬЦЕВОЙ СЕКРЕЦИИ выводится пенициллин (на 80-90 %).
при ПОРАЖЕНИИ различных отделов НЕФРОНА ряд ЛЕКАРСТВЕННЫХ соединений длительно циркулируют в крови и могут не выделяться из организма.
В этих случаях НЕОБХОДИМО изменение дозировок ЛЕКАРСТВЕННЫХ веществ.